Хойт С.К. Генетическая структура европейских ойратских групп по локусам ABO, RH, HP, TF, GC, ACP1, PGM1, ESD, GLO1, SOD-A
// Проблемы этнической истории и культуры тюрко-монгольских народов. Сборник научных трудов. Вып. I. Элиста: КИГИ РАН, 2009. с. 146-183.
текст содержит фрагменты на монгольском, правила чтения тут
Современные европейские ойратские группы, более известные как калмыки, населяют территории Северо-западного Прикаспия, расположенные примерно на 4500 км западнее мест расселения ойратских групп в XVII в. Первые упоминания об ойратах относятся к XIII в. [Сокровенное сказание монголов 1990]. В этот период они локализуются в верховьях Енисея, Приангарье и части Западного Прибайкалья [Эрдниев 1985: 23]. Позже, во времена Чингис-хаана и его преемников, ойратские кочевья передвигаются в Алтайские горы [Далай 1983: 53], где и пребывают до распада империи. В первой половине XVII в. ойраты ставят под контроль значительные территории, расположенные от современной Центральной Монголии до Волги и Дона. Известно, что состав ойратского союза в разные периоды изменялся [Шилэгийн Норбо 1999]. В данный период он включал зvvнгар, дєрвєд, торгууд, хошууд, хойд. В литературе эти группы часто ошибочно именуются племенами, но в XVII в. они являются феодальными владениями. В силу политических и экономических причин этнический состав этих феодальных владений не был стабильным, хотя, по всей видимости, ядро основных родов сохранялось в каждом владении на протяжении всего его существования. Основные группы союза XVII в. легко прослеживаются в составе современных ойрат, немногим отличающемся от средневекового. Желающих ознакомиться с данными по локализации и численности современных ойратских групп мы отсылаем к подробной сводке опубликованной в прошлом году [Хойт 2008].
Рассматриваемая нами европейская ветвь ойрат в XVII в. содержала практически все вышеперечисленные группы, однако соотношение их было неодинаковым. Самым многочисленным, на порядок превышающим другие группы, было торгуудское население. После ухода в 1771 г. большей его части под руководством наместника Убаши на территории бывшего зvvнгарского владения, соотношение торгууд и дєрвєд на европейских территориях стало примерно равным. Хошууд хотя и сохранились до наших дней, однако в силу изначальной немногочисленности были частично растворены в составе торгууд. Остальные, еще более немногочисленные группы, дошли до наших дней лишь в виде родов в составе торгууд, дєрвєд и бузава. Различными комбинациями перечисленных выше этнических групп являлись относительно немногочисленные территориальные группы (уральские, терские, кумские и т.д.), в подавляющем большинстве до наших дней не сохранившиеся. Исключение составляют бузава (бузаава), сложившиеся уже на европейской территории. Эта группа сложилась на Дону из торгууд, дєрвєд и прикочевавших позднее зvvнгар. Таким образом, современные европейские ойраты представлены четырьмя группами: торгууд, дєрвєд, хошууд и бузава. Все вместе, в русскоязычной литературе, они фигурируют под общим этнонимом калмык (калмак, халимаг).
Следует отметить, что исследование истории сложения различных (не только ойратских) групп в разных областях (археология, история, этнография, лингвистика, антропология, генетика) до сих пор протекает параллельно. При этом массивы существующих данных неоднородны, разрозненны и зачастую информация из одной области никак не используется для сопоставления с данными из другой. Что касается проблем генезиса и развития именно ойратских групп, то они до сих пор не разработаны в достаточной мере даже историками. Освещение этих проблем посредством данных этнографии, лингвистики, антропологии и генетики также нельзя назвать удовлетворительным. Между тем генетические, а именно популяционно-генетические методы, традиционно используемые для решения чисто биологических задач, являются одним из инструментов, позволяющим рассмотреть вопросы этногенеза с новой точки зрения и внести вклад в разрешение ряда исторических вопросов.
В данной работе исследуется генетический полиморфизм основных европейских ойратских групп (далее калмыков). Поскольку работа предназначается не столько для антропологов и генетиков, сколько для специалистов из гуманитарных областей, структура ее изложения будет отличаться от традиционной наличием подробных, по возможности, объяснений.
Результаты предыдущих антропометрических исследований [Чебоксаров, 1935; Левин, Трофимова, 1937; Ашилова, 1976] дают достаточно веские причины предполагать, что, несмотря на возможность смешения друг с другом, генетический облик различных ойратских групп в прошлом отличался и, по всей видимости, эти отличия могут быть обнаружены при изучении современного населения.
Напомним, что в основе любой систематики лежит посылка, согласно которой различия либо сходства между группами трактуются как последствия раннего либо позднего, соответственно, их расхождения. При этом следует помнить, что генетическое сходство может объясняться как общей историей групп, так и давлением отбора, поэтому при анализе сходства и различия между популяциями обычно стараются использовать как можно большее число признаков. Большее число генов, вовлеченных в анализ, уменьшает вероятность того, что все они находятся под одинаковым давлением отбора [Фогель, Мотульски 1990: 364]. На практике, из-за сложности изучения генетического полиморфизма (от сбора образцов крови до дорогостоящего анализа), данные по распространению генов обычно охватывают куда меньшее число генов, чем нам того хотелось бы.
В данной работе представлены результаты нашего популяционно-генетического исследования 1995-1997 гг., в котором на основании частот аллелей ряда белковых систем генетических маркеров (ABO, RH, HP, TF, GC, ACP1, PGM1, ESD, GLO1, SOD-A) устанавливается место калмыков в системе мирового распределения частот генов и исследуется генетическая дифференциация основных калмыцких групп. Основные результаты исследования были опубликованы ранее [Ovtchinnikova et al., 1997; Galushkin, 1998; Galushkin et al., 2001], однако в силу специфики популяционно-генетических статей, основной упор в них был сделан на представлении именно результатов, их интерпретации и анализу отводилась довольно скромная роль. Эта работа отличается от предыдущих тем, что, помимо частот генов, в ней представляются новые, расширенные результаты, основанные на большем количестве аналитических процедур, в которые были включены данные по калмыцким группам и ряду этнических групп Евразии (отсутствовавших в предыдущем анализе), кроме того, интерпретация этих результатов проведена на качественно ином уровне с привлечением различного рода исторических и лингвистических данных.
Материалы и методы
Сбор образцов. Материал – 145 образцов крови был собран в сентябре 1995 г. в г. Элисте, на базе Калмыцкой республиканской станции переливания крови. Кровь отбиралась у доноров, не связанных прямым родством, идентифицировавших себя калмыками, живущих в г. Элисте и в районах республики. Помимо этнической идентификации (на разных уровнях), с помощью интервью выяснялся ряд других сведений, в том числе наличие в родословных возможных иноэтничных включений.
На основании результатов интервью данные по генетическим частотам рассчитывались для нескольких групп. В общую выборку «калмыки» попадали результаты всех людей, в том числе метисов в первом поколении (4 чел.), которые идентифицировали себя как калмыков. Помимо метисов, в эту же общую выборку были включены данные еще 2 чел. Один из них оказался хошуудом, другой, назвав себя калмыком, отказался далее конкретизировать свою этническую принадлежность. Для отдельных групп (торгууд, дєрвєд и бузава) генетические частоты рассчитывались как для индивидов, принадлежащих группе по линии отца и матери по крайней мере до второго поколения (т.е. по 4-м предковым линиям), так и относящихся к группе по линии отца, без учета материнских линий. В первом случае группы условно назывались «несмешанными», т.е. «несмешанные торгууд», «несмешанные дєрвєд», «несмешанные бузава». Во втором случае – просто «торгууд», «дєрвєд», «бузава».
Лабораторные процедуры. Образцы обрабатывались c мая 1996 г. по май 1997 г. на базе и по методикам, используемым в лаборатории экологической генетики Института клинической генетики Медико-генетического научного центра Российской академии медицинских наук. Группы крови ABO и RH были типированы стандартными серологическими методами [Алексеева и др. 1970]. Генотипы гаптоглобина (HP) были типированы с помощью обычного электрофореза на вертикальных полиакриламидных гелях с небольшими количествами гемолизата, добавленного к образцам сыворотки [Спицын 1985]. Трансферрин (TF) и группоспецифический компонент (GC) типировались изоэлектрическим фокусированием в том же полиакриламидном геле в присутствии иммобилинов методом Gorg et al. [1983]. GC обнаруживался методом изоэлектрического фокусирования (ИЭФ), описанного Спицыным и Титенко [1990]. Кроме того, методом ИЭФ были типированы эритроцитарные ферменты: кислая фосфатаза 1 (ACP1) на агарозном геле методом Miller et al. [1987]; эстераза D (ESD) на полиакриламидном геле методом Divall [1994]; а также фосфоглюкомутаза-1 (PGM1) и супероксиддисмутаза-A (SOD-A) на полиакриламидных гелях методом Goedde et al. [1981].
Группы сравнения. Для сравнения частот генов европейских ойрат (калмыков) с частотами генов других этнических групп Евразии было бы интересно взять современные группы, близкие к исходным предковым популяциям, и группы, с которыми ойраты взаимодействовали в течение различных периодов своей истории. К первой группе можно причислить часть тунгусо-маньчжуроязычных, все монголоязычные и большую часть тюркоязычных народов Сибири. Для полноты картины желательно ввести в анализ данные по корейцам и японцам. Ко второй группе можно отнести популяции современной КНР (прежде всего, конечно, хань, и, возможно, южнокитайские группы), тюркоязычные популяции степного пояса, популяции Северного Кавказа, русских. В идеале для выяснения размаха евроазиатской изменчивости можно было бы также взять некоторые группы Западной Европы (например, немцев, французов), но в силу разных причин от западноевропейских популяций мы отказались.
Для основной части групп сравнения использовалась генетические частоты из базы данных «Gene pool» (разработана под руководством Е.В. Балановской; программное обеспечение А.Н. Ивлева, В.Е. Папкова на базе лабораторий генетики человека ИОГен РАН, под рук. проф. Ю.Г. Рычкова и генетической эпидемиологии ИКГ РАМН, под рук. чл.-корр. Е.Р. Гинтера). Данные базы «Gene pool» представлены в виде обобщенных средних значений генных частот. Они были дополнены информацией из сводки «Генофонд и геногеография населения» [здесь и далее – ГГН, 2000], являющейся, в сущности, альтернативной опубликованной базой данных. Кроме этого, для различных монголоидных групп привлекалась информация разбросанная по отдельным статьям.
Таблица 1. Группы сравнения.
| обозначение на рисунках | локализация популяций | источник |
| якуты | Gene pool | |
| яуты Кюсюр | Якутская АССР, Булунский р-н, пос. Кюсюр, п.г.т. Тикси | ГГН, 2000 |
| якуты Ессей | Якутская АССР, Эвенкийский авт. окр. Илимпийский р-н с. Ессей | ГГН, 2000 |
| японцы | Япония, основные острова (Japan main islands) | Omoto et al., 1973 |
| корейцы | Gene pool | |
| корейцы Каннын | Южная Корея, г. Каннын (Kangreung) | Benkman et al., 1989 |
| корейцы Кванджу | Южная Корея, г. Кванджу (Kwangju) | Benkman et al., 1989 |
| корейцы Чеджу | Южная Корея, г. Чеджу (Cheju) | Benkman et al., 1989 |
| корейцы Тэджон | Южная Корея, г. Тэджон (Taejon) | Benkman et al., 1989 |
| корейцы Сеул | Южная Корея, г. Сеул (Seoul) | Benkman et al., 1989 |
| корейцы Гирин | Гирин КНР (Jilin) | Du, Chen, 1989 |
| эвены Анюйск | Чукотский авт. окр. Билибинский р-н с. Анюйск | ГГН, 2000 |
| эвены Омолон | Чукотский авт. окр. Билибинский р-н с. Омолон | ГГН, 2000 |
| эвенки зап. | Gene pool | |
| эвенки зап. Байкит | Эвенкийский авт. окр. Байкитский р-н села Байкит и Суринда | ГГН, 2000 |
| эвенки зап. Юкта | Эвенкийский авт. окр. Илимпийский р-н поселки Юкта и Кислокан | ГГН, 2000 |
| эвенки зап. Нидым | Эвенкийский авт. окр. Илимпийский р-н поселки Нидым и Учами | ГГН, 2000 |
| эвенки зап. Чиринда | Эвенкийский авт. окр. Илимпийский р-н с. Чиринда | ГГН, 2000 |
| эвенки зап. Эконда | Эвенкийский авт. окр. Илимпийский р-н с. Эконда | ГГН, 2000 |
| эвенки зап. Кюсюр | Якутская АССР Булунский р-н пос. Кюсюр, п.г.т. Тикси | ГГН, 2000 |
| эвенки вост. | Gene pool | |
| эвенки АРВМ | АРВМ КНР (Inner Mongolia) | Du, Chen, 1989 |
| орочоны АРВМ, Хэйлунцзян | АРВМ, Хэйлунцзян КНР (Inner Mongolia, Heilongjiang) | Du, Chen, 1989 |
| монголы МР | Монгольская республика г. Улаанбаатар | Батсуурь и др., 1991 |
| монголы АРВМ | АРВМ КНР (Inner Mongolia) | Du, Chen, 1989 |
| дауры Хэйлунцзян | Хэйлунцзян КНР (Heilongjiang) | Du, Chen, 1989 |
| буряты | Gene pool | |
| буряты Судунтуй | Агинский Бурятский авт. окр., с. Судунтуй | Novoradovsky et al., 1993 |
| буряты Сахюрта | Иркутская обл., Ольхонский р-н, м.р.с. Сахюрта (?Агинский Бурятский авт. окр., с. Сахюрта) | Novoradovsky et al., 1993 |
| буряты Усть-Орда | Усть-Ордынский Бурятский авт. окр., п.г.т. Усть-Орда | Novoradovsky et al., 1993 |
| буряты Ольхон | о. Ольхон | Novoradovsky et al., 1993 |
| буряты Могсохон | Республика Бурятия, Кижигинский р-н, с. Могсохон | Novoradovsky et al., 1993 |
| хань Сев.Кит. | Северный Китай (North China) | Du, Chen, 1989 |
| тайваньцы Тайчунг | о. Тайвань г. Тайчунг | Lin Jing Y., 1975 |
| чжуан Гуанси | Гуанси КНР (Guangxi) | Du, Chen, 1989 |
| дун Гуанси | Гуанси КНР (Guangxi) | Du, Chen, 1989 |
| уйгуры | Gene pool | |
| уйгуры СУАР | СУАР КНР (Xinjiang) | Du, Chen, 1989 |
| алтайцы северные | Gene pool | |
| алтайцы южные | Gene pool | |
| шорцы | Gene pool | |
| тувинцы | Gene pool | |
| тоджинцы | Gene pool | |
| тофалары | Gene pool | |
| хакасы | Gene pool | |
| киргизы | Gene pool | |
| киргизы Фрунзе | Киргизская ССР г. Фрунзе | ГГН, 2000 |
| казахи | Gene pool | |
| казахи зап. | Западная этноисторическая область | ГГН, 2000 |
| казахи сев.-цент. | Северо-Центральная этноисторическая область | ГГН, 2000 |
| казахи юж.-вост. | Южно-Восточная этноисторическая область | ГГН, 2000 |
| туркмены | Gene pool | |
| татары | Gene pool | |
| башкиры | РСФСР | ГГН, 2000 |
| ногайцы Дагестан | Дагестанская АССР | ГГН, 2000 |
| адыгейцы | Gene pool | |
| кабардинцы | Gene pool | |
| черкесы | Gene pool | |
| кумыки | Gene pool | |
| даргинцы | Gene pool | |
| чеченцы | Gene pool | |
| русские | Gene pool |
Отметим, что размеры выборок колебались не только для отдельных групп, но и для частот различных генов внутри одной группы. Разброс минимальных и максимальных выборок колебался от десятков человек до нескольких тысяч. Также стоит отметить, что лишь для меньшей части из всех приведенных групп сравнения имелись данные по тем же системам, что и у изученных нами калмыцких популяций. В большинстве случаев набор изученных систем совпадал лишь отчасти.
Аналитические процедуры. Для каждой выделенной нами группы определялась величина наблюдаемой и теоретической гетерозиготности. Средняя гетерозиготность находилась как средняя арифметическая по 10 локусам (АВО, RH, TF, GC, HP, ACP1, PGM1, SOD A, GLO1, ESD) [Животовский 1991]. Генетические расстояния между исследуемыми и другими группами Евразии вычислялись по методу Nei [1972]. На основании полученной матрицы генетических расстояний проводился кластерный анализ и многомерное шкалирование.
Кластерный анализ на основе данных о взаимных расстояниях с применением относительно несложных вычислений жестко и достаточно однозначно выделяет группы таксономических единиц, расстояния между которыми относительно невелики. Визуально ход анализа и его результаты обычно представляются в виде дендрограммы. Вид этой дендрограммы часто может существенно зависеть от выборочных случайностей и от примененной процедуры выявления кластеров.
В отличие от кластерного анализа, результат многомерного шкалирования представляет все расстояния между исследуемыми единицами в пространстве 2 или 3 измерений. Многомерное шкалирование показывает наиболее оптимальное взаимное положение популяций в пространстве комплекса изучаемых признаков. При этом суждение о наблюдаемой таксономической структуре производится визуально, и в этом отношении многомерное шкалирование родственно каноническому или компонентному анализам.
Цель многомерного шкалирования достигается обычно в ходе получения при вычислениях ряда последовательных приближений набора наблюдаемых на графике расстояний к эмпирически вычисленным. Процесс завершается при достижении минимально возможных суммарных различий этих двух типов расстояний между всеми парами рассматриваемых единиц. Наилучшие результаты обычно получаются с применением так называемого неметрического шкалирования, которое накладывает меньше строгих ограничений на соотношения наблюдаемых и эмпирических расстояний. Соответствие эмпирических и наглядно видимых на графике расстояний обычно представляется в виде так называемой диаграммы Шепарда. На ней приводится корреляционное поле для значений этих двух видов расстояний. Хорошему результату соответствует вытянутая вдоль диагонали диаграммы картина этого корреляционного поля.
Степень суммарных различий двух типов расстояний описывается так называемой величиной стресса. Многомерное шкалирование считается успешным, если стресс имеет допустимо малую величину (обычно – меньше 0,10 или 0,15). Таким образом, стресс является показателем качества многомерного шкалирования [Терехина 1986; Дейвисон 1988].
Вполне понятное желание сравнить полученные частоты генов с частотами генов как можно большего числа популяций столкнулось с упомянутой выше неравномерной изученностью различных популяций. Даже с использованием баз данных мы не располагали информацией по всему спектру маркеров для всех интересующих нас групп. В силу этого факта мы были поставлены перед выбором: сравнить наши данные с данными по максимальному набору локусов для минимального числа групп либо сравнить их с данными по минимальному набору локусов для максимального числа групп. В итоге был проделан ряд анализов, результаты которых иллюстрируют оба случая, а также плавный переход между ними. Помимо этого, увеличению числа анализов способствовало еще несколько факторов, подробно разъясняемых ниже.
Переход от анализа «много популяций – мало маркеров» к анализу «мало популяций – много маркеров» осуществлялся увеличением числа систем маркеров. Добавление в анализ каждой следующей системы маркеров осуществлялось на основании наибольшей встречаемости данных по ней среди популяций. Иногда разные системы могли иметь одинаковую встречаемость, в этих случаях производилось несколько анализов с одинаковым числом систем маркеров, но разным их набором.
Механическое добавление в анализ систем маркеров дополнялось другим подходом, когда мы делали приоритетным не набор систем маркеров, а набор тех или иных интересующих нас популяций. Это также увеличило количество вариантов с одинаковым числом систем, но разным их набором.
В силу того, что в анализ последовательно вводились не только частоты генов по калмыкам в целом, но и по торгууд, дєрвєд и бузава, которые в свою очередь подразделялись на группы в целом и «несмешанные» группы, а также в силу того, что данные по группам сравнения также вводились как общие средние частоты генов и частоты по их локальным популяциям, число всех возможных вариантов подсчета резко выросло.
В некоторых случаях группы сравнения формировались несколькими способами (все возможные, с отсечением явно европеоидных, тюрко- и монголоязычные, только монголоязычные), что также увеличило чисто вариантов подсчета.
Последний фактор, увеличивший число вариантов подсчета, заключается в том, что частота одних аллелей обусловлена частотой других. Например: если в двуаллельной системе один аллель будет представлен с частотой 60%, то второй – обязательно 40%; в трехаллельной системе, если один аллель будет иметь частоту 40%, второй – 40%, то третий – обязательно 20%... Таким образом, для сравнения популяций по двуаллельной системе достаточно иметь данные по частоте одного аллеля, по трехаллельной системе – по частоте двух аллелей... Данные по частоте второго и третьего аллелей для дву- и трехаллельных систем соответственно в математике носят название избыточной информации. При решении поставленной задачи мы по нашему выбору можем использовать или не использовать ее. Для подавляющего большинства случаев были просчитаны оба варианта.
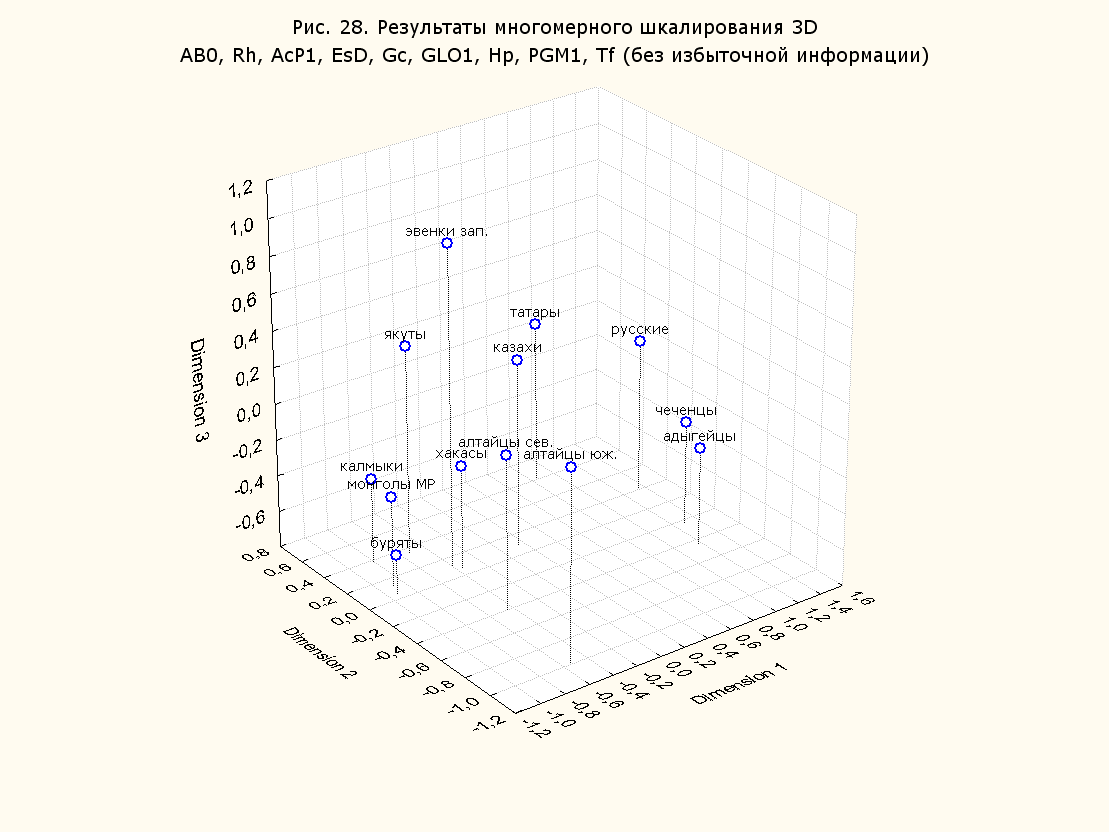
Как видим, различные факторы значительно увеличили число возможных вариантов анализа. Из всего этого множества вариантов было подсчитано порядка 150, однако в силу ограниченности объема публикации, ниже будут представлены лишь некоторые из них. По этой же причине мы представляем генетические расстояния только в пространстве двух переменных (2D). Варианты в пространстве трех переменных (3D, за исключением рис. 28, 29), и диаграммы Шепарда для каждого случая будут опущены.
Результаты
В отличие от предыдущей работы [Galushkin et al. 2001], где были приведены частоты аллелей серологических и биохимических систем, а также частоты их фенотипов, в данном случае мы привели только частоты аллелей (таб. 2). Еще одним отличием от предыдущей работы является то, что эти частоты приведены как для групп в целом, так и для «несмешанных» групп.
По большинству систем приведены данные для 145 чел. По иммунологическим маркерам АВО и RH – данные для 158 чел. По системе НР у 2 чел. был обнаружен фенотип 0, происхождение которого трудно объяснить. Наличие этого фенотипа может объясняться либо мутацией, либо грубой патологией. Поэтому по данной системе эти индивиды не учитывались, данные приведены для 143 чел.
Значения гетерозиготности, наблюдаемой и ожидаемой, во всех вышеперечисленных группах представлены в таб. 3.
Таблица 2. Частоты аллелей.
| система, аллели | калмыки | торгууд | дєрвєд | бузава | торгууд несмешанные | дєрвєд несмешанные | бузава несмешанные |
| RH (N) | 158 | 63 | 64 | 21 | 49 | 48 | 13 |
| *D | .9204 | 1,0 | 1,0 | .7818 | 1,0 | 1,0 | 1,0 |
| *d | .0796 | 0,0 | 0,0 | .2182 | 0,0 | 0,0 | 0,0 |
| AB0 (N) | 158 | 63 | 64 | 21 | 49 | 48 | 13 |
| *A | .1975 | .2489 | .1547 | .2026 | .2563 | .1103 | .1663 |
| *B | .2733 | .2713 | .2976 | .2319 | .2149 | .2785 | .2636 |
| *0 | .5292 | .4798 | .5477 | .5655 | .5288 | .6111 | .5701 |
| TF (N) | 145 | 61 | 59 | 19 | 48 | 44 | 11 |
| *C1 | .7724 | .7705 | .7881 | .7632 | .7604 | .8068 | .7273 |
| *C2 | .1793 | .1557 | .1864 | .2368 | .1563 | .1705 | .2727 |
| *C3 | .0138 | 0.0 | .0254 | 0.0 | .0833 | .0227 | 0.0 |
| *D | .0345 | .0738 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| GC (N) | 145 | 61 | 59 | 19 | 48 | 44 | 11 |
| *1F | .4759 | .5410 | .4407 | .3421 | .5104 | .3977 | .1364 |
| *1S | .2517 | .2951 | .2034 | .3158 | .3333 | .1932 | .5000 |
| *2 | .2724 | .1639 | .3559 | .3421 | .1563 | .4091 | .3636 |
| HP (N) | 143 | 59 | 59 | 19 | 47 | 44 | 11 |
| *1 | .3182 | .3644 | .2373 | .3947 | .3511 | .2159 | .3636 |
| *2 | .6818 | .6356 | .7627 | .6053 | .6489 | .7841 | .6364 |
| ACP1 (N) | 145 | 61 | 59 | 19 | 48 | 44 | 11 |
| *A | .2621 | .2459 | .2797 | .2368 | .2396 | .2727 | .2727 |
| *B | .7207 | .7459 | .6864 | .7632 | .7604 | .6932 | .7273 |
| *C | .0172 | .0082 | .0339 | 0.0 | 0.0 | .0341 | 0.0 |
| PGM1 (N) | 145 | 61 | 59 | 19 | 48 | 44 | 11 |
| *1+ | .6828 | .6967 | .6610 | .6842 | .6771 | .6591 | .5909 |
| *1- | .0897 | .1066 | .0763 | .0789 | .1250 | .0795 | .0909 |
| *2+ | .1897 | .1639 | .2203 | .1842 | .1667 | .2273 | .2727 |
| *2- | .0379 | .0328 | .0424 | .0526 | .0313 | .0341 | .0455 |
| ESD (N) | 145 | 61 | 59 | 19 | 48 | 44 | 11 |
| *1 | .7897 | .7459 | .8136 | .8947 | .7292 | .8068 | .9091 |
| *2 | .2000 | .2459 | .1695 | .1053 | .2708 | .1705 | .0909 |
| *5 | .0034 | .0082 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| *7 | .0069 | 0.0 | .0169 | 0.0 | 0.0 | .0227 | 0.0 |
| SOD A (N) | 145 | 61 | 59 | 19 | 48 | 44 | 11 |
| *1 | .9931 | 1.0 | .9831 | 1.0 | 1.0 | .9773 | 1.0 |
| *2 | .0069 | 0.0 | .0169 | 0.0 | 0.0 | .0227 | 0.0 |
| GLO1 (N) | 145 | 61 | 59 | 19 | 48 | 44 | 11 |
| *1 | .1690 | .1721 | .1525 | .1842 | .1875 | .1591 | .1818 |
| *2 | .8310 | .8279 | .8475 | .8158 | .8125 | .8409 | .8182 |
| локус | калмыки | торгууд | дєрвєд | бузава | торгууд несмешанные | дєрвєд несмешанные | бузава несмешанные | |
| ACP1 | набл. | .4414 | .4098 | .4746 | .3684 | .3542 | .4318 | .3636 |
| ожид. | .4116 | .3831 | .4494 | .3615 | .3644 | .4440 | .3967 | |
| ESD | набл. | .3655 | .4426 | .3051 | .2105 | .4583 | .2955 | .1818 |
| ожид. | .3364 | .3831 | .3091 | .1884 | .3950 | .3195 | .1653 | |
| GLO1 | набл. | .3379 | .3443 | .3051 | .3684 | .3750 | .3182 | .3636 |
| ожид. | .2808 | .2850 | .2585 | .3006 | .3047 | .2676 | .2975 | |
| GC | набл. | .5517 | .5738 | .5932 | .3684 | .5833 | .6136 | .1818 |
| ожид. | .6360 | .5934 | .6377 | .6662 | .6039 | .6371 | .5992 | |
| HP | набл. | .4965 | .5254 | .4068 | .6842 | .5745 | .3864 | .7273 |
| ожид. | .4339 | .4632 | .3620 | .4778 | .4556 | .3386 | .4628 | |
| PGM1 | набл. | .4897 | .4590 | .5424 | .4211 | .4792 | .5227 | .4545 |
| ожид. | .4884 | .4753 | .5069 | .4889 | .4972 | .5065 | .5661 | |
| SOD A | набл. | .0138 | .0339 | .0455 | ||||
| ожид. | .0137 | .0333 | .0444 | |||||
| TF | набл. | .3517 | .3443 | .3051 | .4737 | .3750 | .2727 | .5455 |
| ожид. | .3698 | .3766 | .3434 | .3615 | .3904 | .3195 | .3967 | |
| средняя | набл. | .3810 | .3874 | .3708 | .3618 | .3999 | .3608 | .3523 |
| ожид. | .3713 | .3670 | .3625 | .3556 | .3764 | .3597 | .3605 | |
Общая характеристика частот генов. В целом калмыки могут быть охарактеризованы высокой частотой аллеля ABO*B. По трехаллельной системе групп крови ABO в трех калмыцких группах наблюдалось следующее отношение частот: О>В>A. Высокая частота группы крови В характерна для монгольских и североазиатских популяций. Положительный резус антиген (RH*D) выявился у всех калмыков, за исключением одного индивидуума. Высокая частота аллеля GC*1F у калмыков сильно походит на частоту, наблюдаемую среди бурят [Novoradovsky et al. 1993]. Распределение частот аллелей PGM1 у калмыков почти идентично частотам, наблюдаемым среди бурят. Характер этого распределения заметно отличается от распределения у эвенков, которые имеют значительно более высокие частоты PGM*1+ и более низкие частоты PGM*1- и 2- [Crawford et al. 2001]. Между калмыцкими группами имеются значительные различия в частотах аллелей ESD. Несмотря на эти различия, в целом калмыки могут быть охарактеризованы наличием высокой частоты аллеля ESD*2 (приблизительно 20%), характерного для азиатских популяций. Редкие аллели ESD*5 и ESD*7 наблюдались в двух основных калмыцких группах. Аллель GLO1*2 достигает самой высокой частоты 80-85% в центральноазиатских популяциях и приблизительно 40% в европейских [Спицын и др. 1994]. Частота аллеля GLO1*2 у калмыков (83%) находится в пределах центральноазиатских показателей. Калмыки отличаются от большинства азиатских групп высокой частотой аллеля SOD-A*1. Обычно редкий аллель SOD-A*2 этого фермента был найден с частотой 0.017% у дєрвєд.
Частоты аллелей, показанные в таб. 2, ясно свидетельствуют, что современные калмыки – азиатская популяция. Почти полное отсутствие резус отрицательного фенотипа и высоких частот аллелей: ABO*B (27,3%); TF*C2 (17,9%); GC*1F (47,6%); ESD*2 (20%); GLO1*2 (83,1%), указывают на то, что калмыки – представители Центральной Азии. Они имеют значительную близость к монголам Монголии [Батсуурь и др. 1991] и бурятам России [Novoradovsky et al. 1993], достоверно отличаясь от европейских популяций.
Характеристика частот генов групп. Калмыцкие группы генетически отличаются друг от друга. В частности, бузава и дєрвєд имеют более низкие частоты ABO*B, GC*1F и ESD*2 аллелей, используемых для определения центральноазиатских популяций. Аллели, характеризующие европеоидов TF*C3 и ACP1*C с частотой 2,5% и 3,4%, соответственно, были обнаружены среди дєрвєд. Аллель SOD-A*2, наибольшие частоты которого установлены у населения Восточной Скандинавии и некоторых народов Северной Евразии, был также найден у дєрвєд (2 чел., частота 0.017%). Единственный резус отрицательный индивидуум был найден среди бузава. Фенотип HP 0 (агаптоглобин), был обнаружен у двух торгууд.
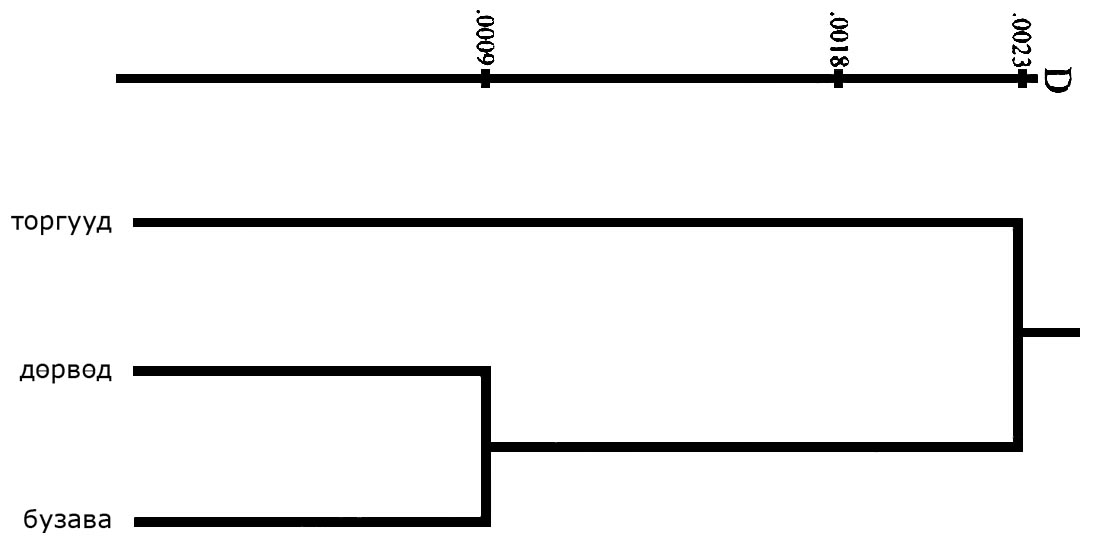
Генетические расстояния между несмешанными торгууд, дєрвєд и бузава, полученные по методу Nei [1972], на основании данных по 10 локусам (ABO, RH, ACP1, PGM1, TF, GC, HP, GLO1, ESD, SOD-A) представлены в таб. 4.
Таблица 4. Генетические расстояния между тремя калмыцкими группами, рассчитанные по методу Nei [1972].
| группа | торгууд | дєрвєд | бузава |
| торгууд | 0,00213 | 0,00207 | |
| дєрвєд | 0,00030 |
Дендрограмма, демонстрирующая относительную близость трех несмешанных калмыцких групп, представлена на рис. 1. Она основана на генетических расстояниях Nei [1972], вычисленных по частотам тех же 10 генетических локусов и показывает, что дєрвєд и бузава генетически намного более близки друг к другу, чем каждая из них торгуудам.
Рисунок 1. Результаты кластерного анализа для трех калмыцких групп, основанные на генетических расстояниях, полученных по методу Nei [1972].
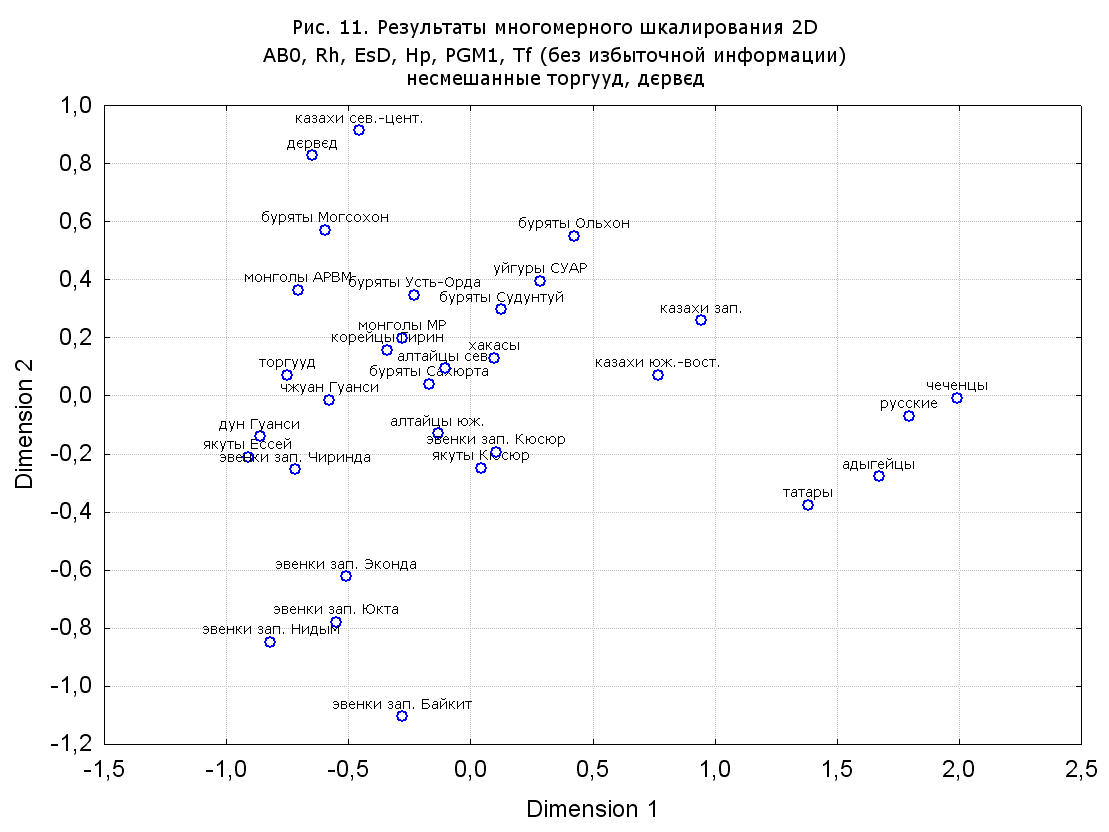
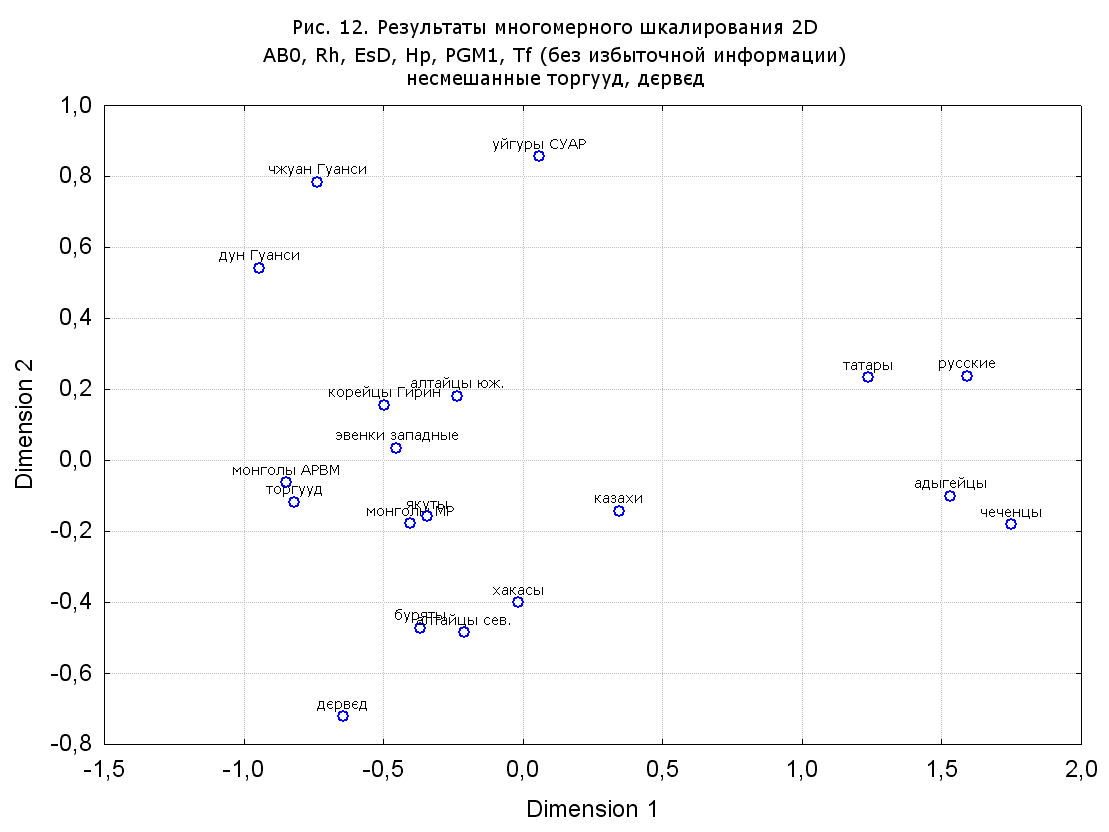
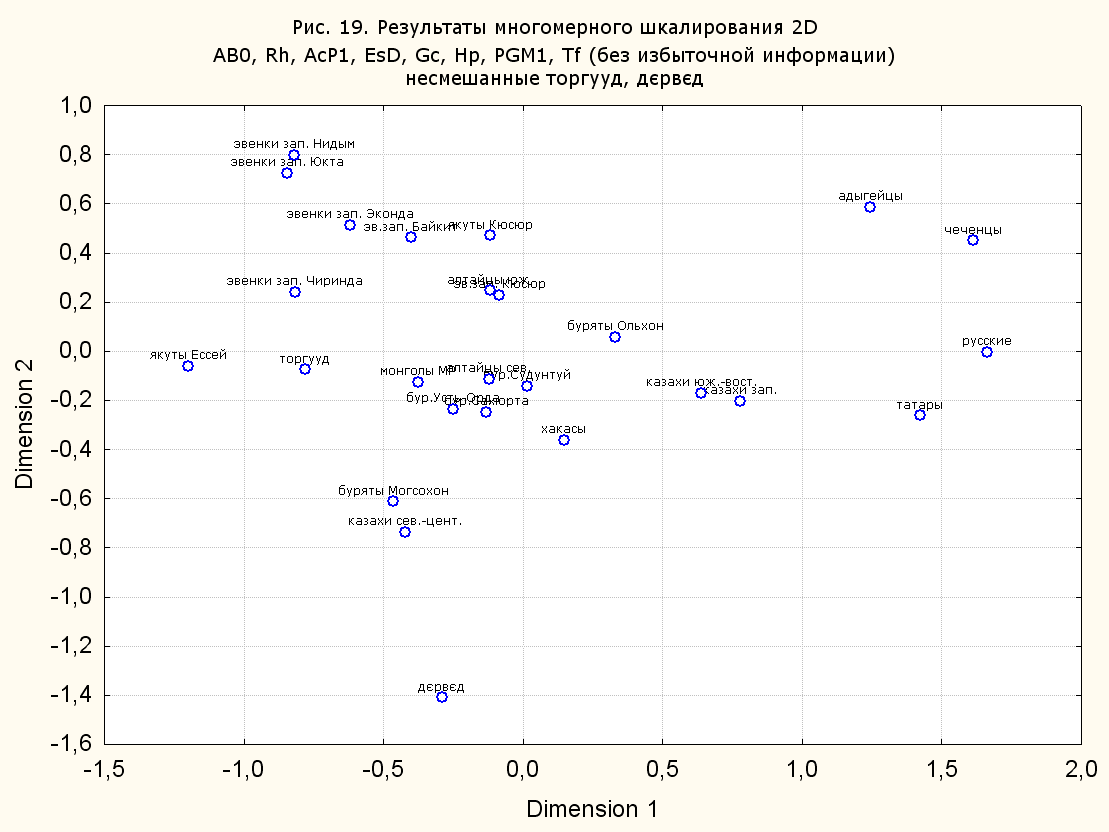
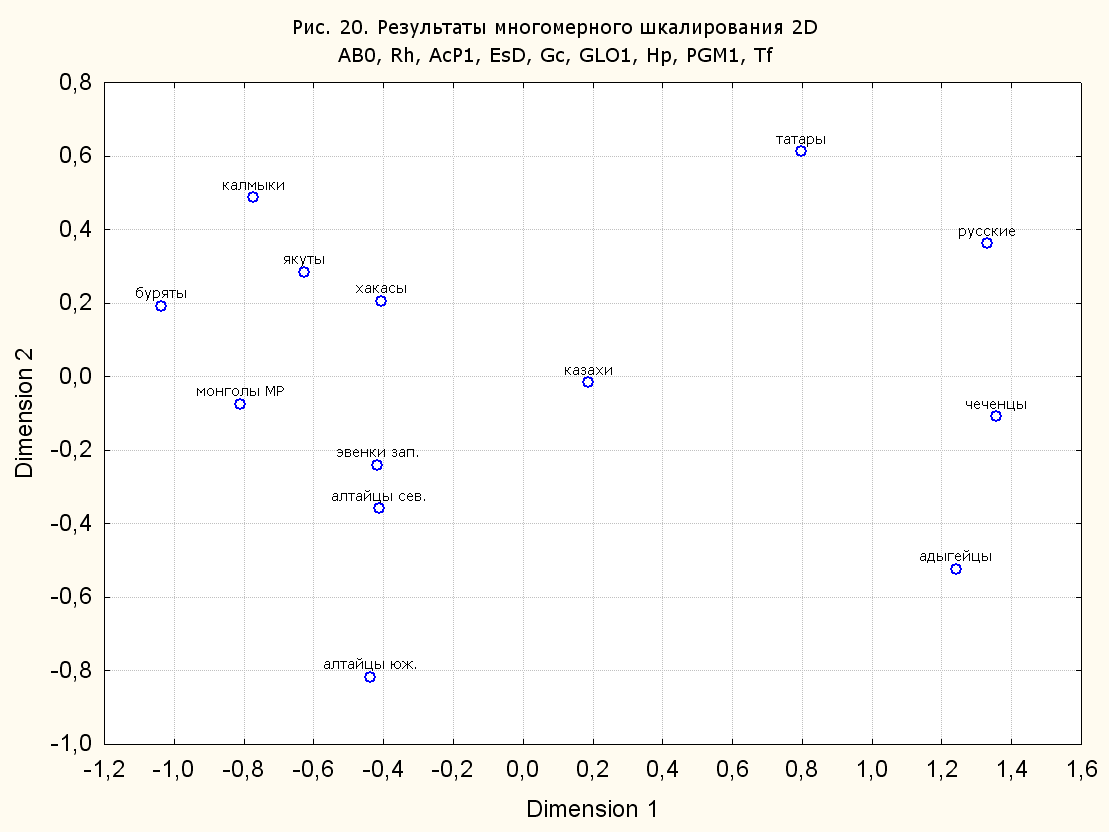
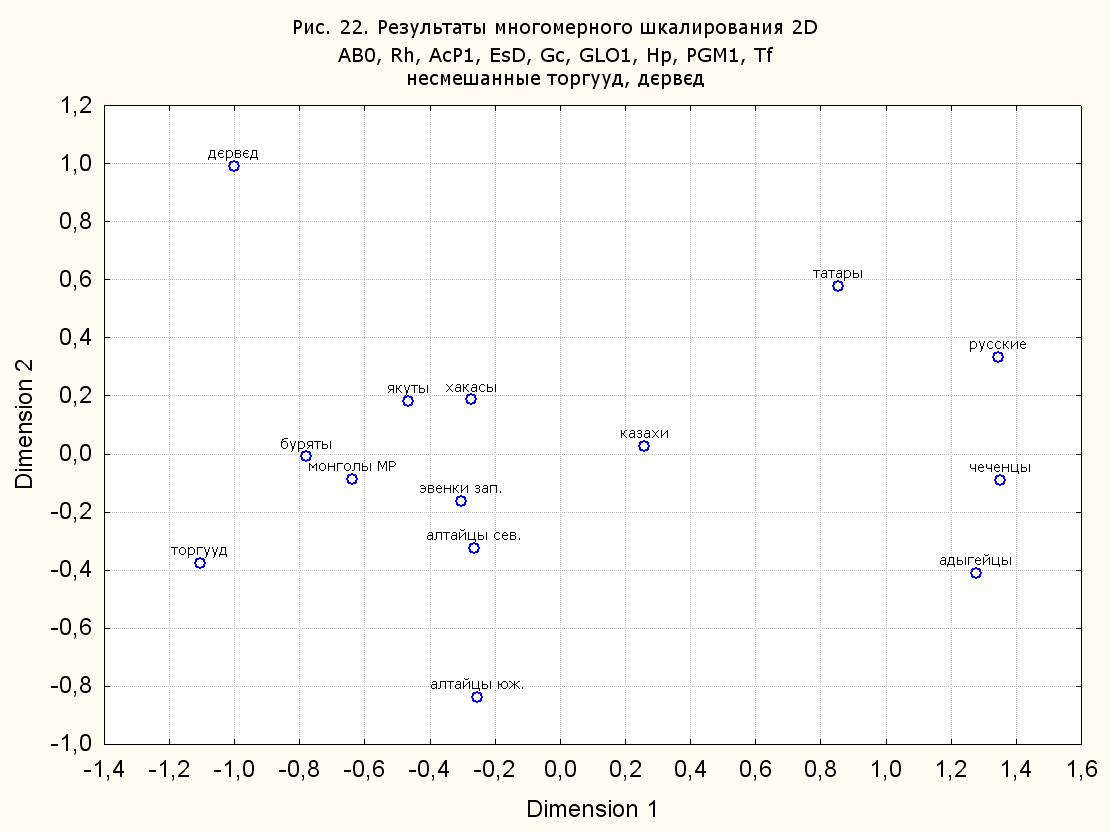
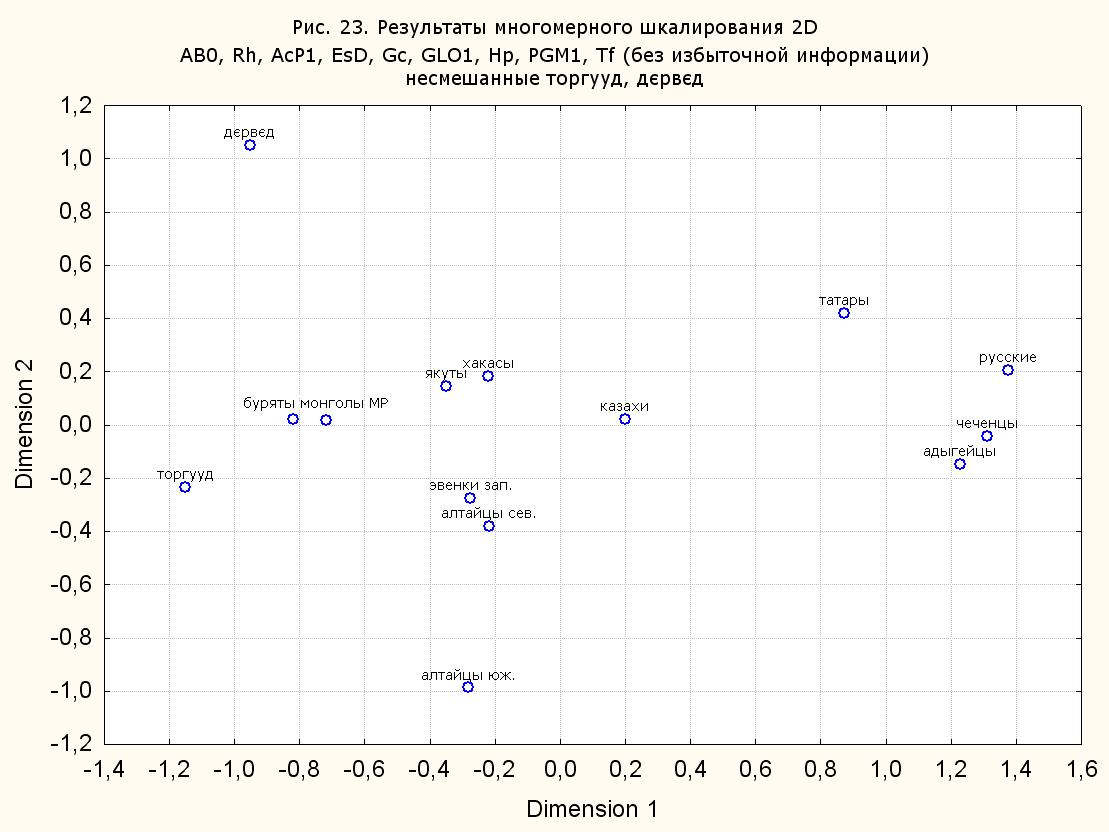
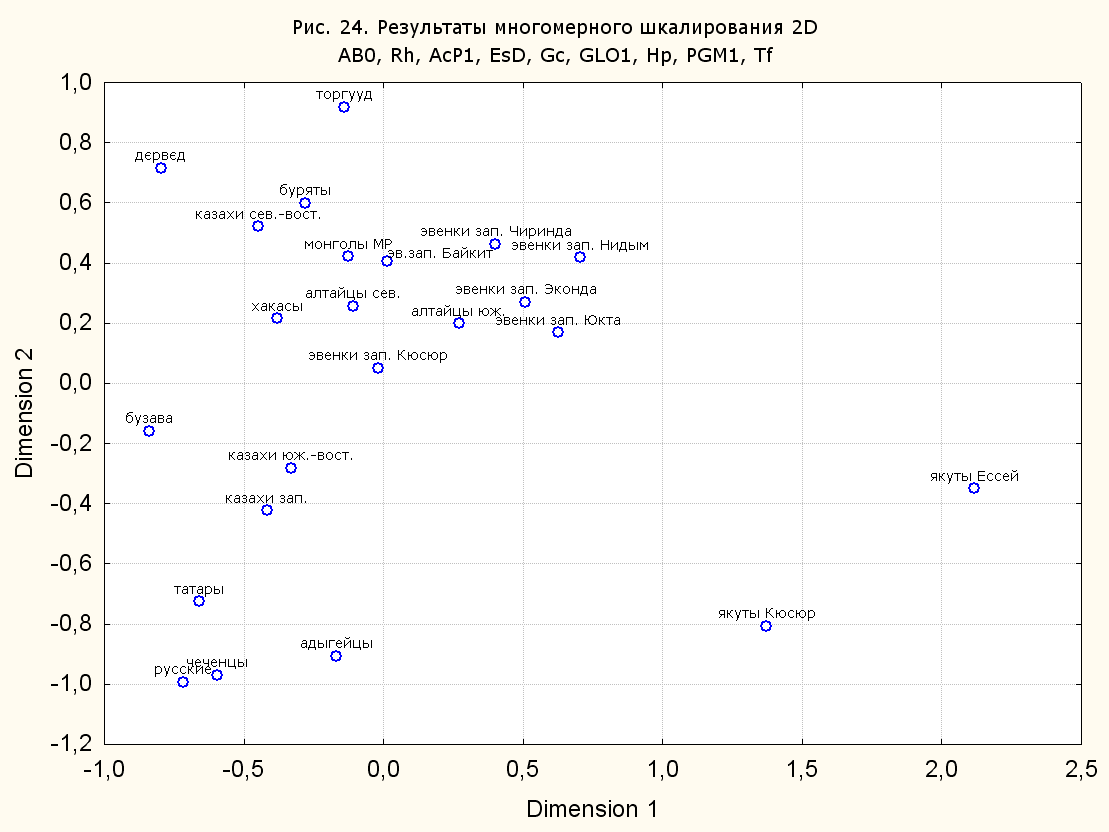
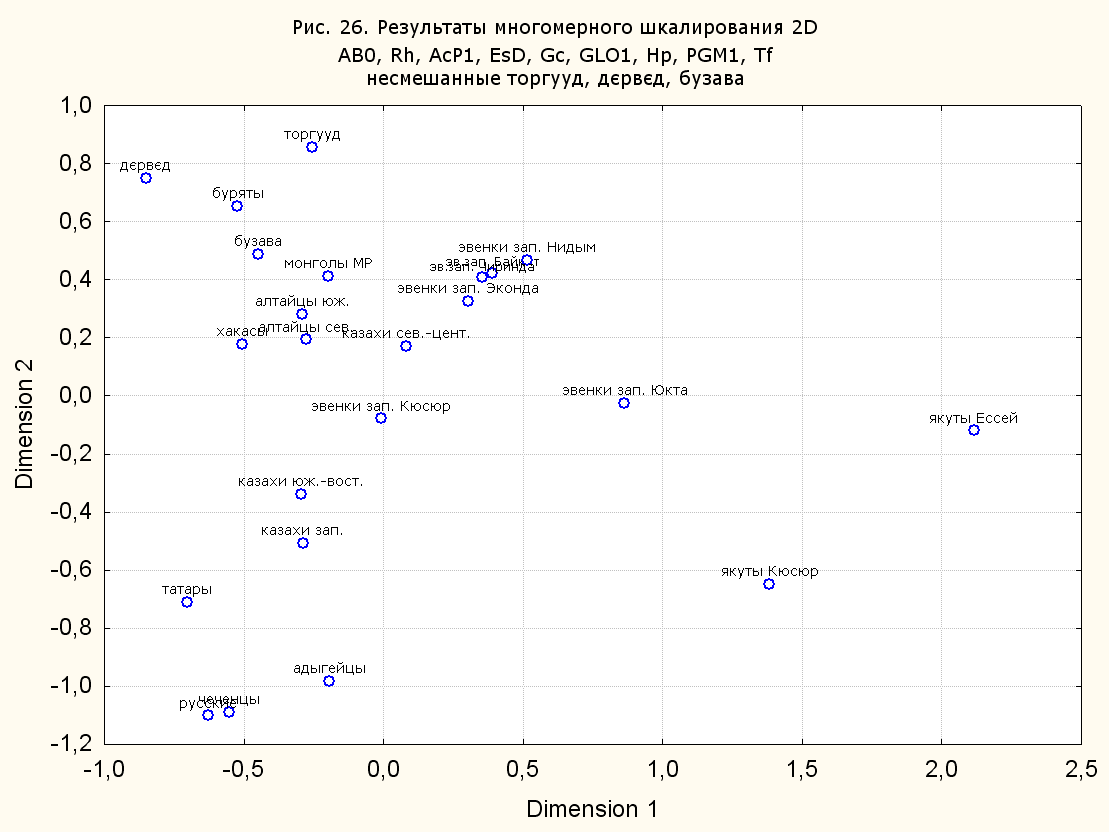
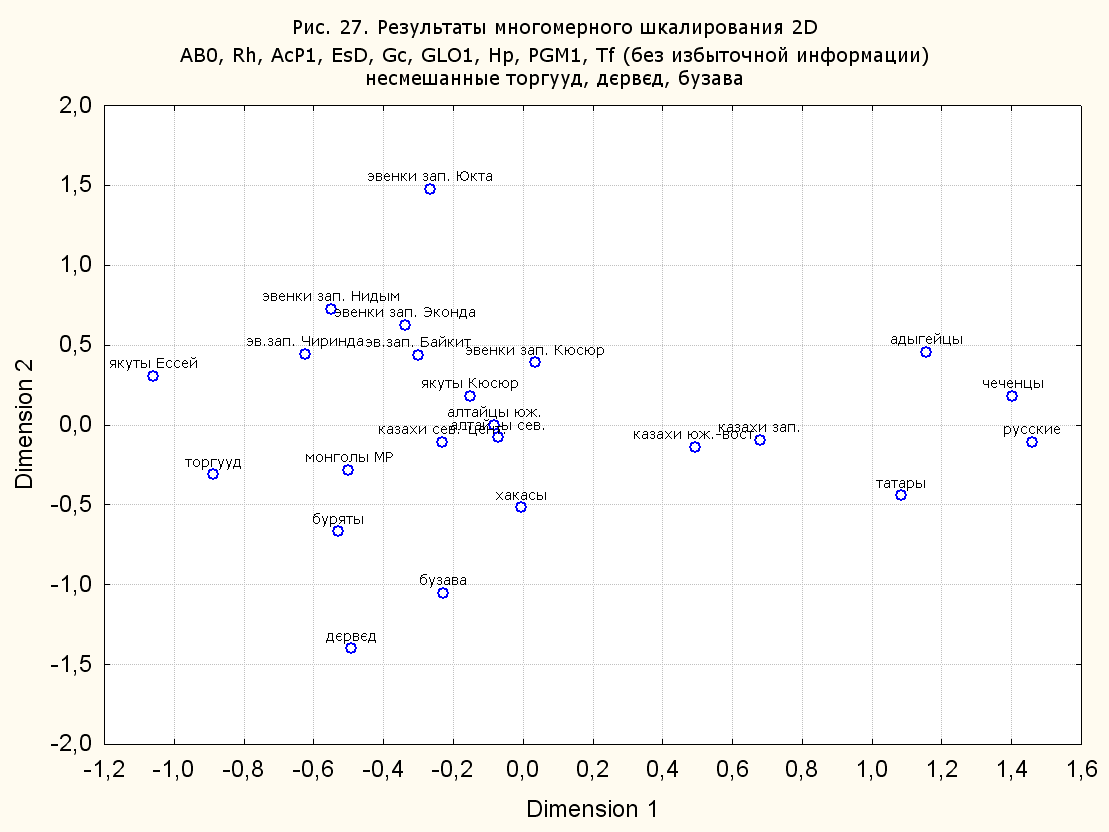
Результаты многомерного шкалирования. Результаты многомерного шкалирования представлены на серии рисунков (рис. 2-29). На основании разных вариантов подсчета по 9 системам мы пришли к выводу, что легче интерпретируемые результаты получаются на основании использования частот генов без избыточной информации. Часть демонстрирующих эту разницу вариантов приведена на рис. 20-27. При делении калмыков на отдельные группы, наиболее выраженные тенденции наблюдаются при использовании частот несмешанных групп. Поэтому, учитывая ограниченность публикации, в большинстве случаев мы привели результаты для частот генов без избыточной информации и для несмешанных групп.
Все результаты многомерного шкалирования, полученные на основе средних частот генов по калмыкам в целом (рис. 2-5, 7, 9-10, 13, 15, 17, 20, 21, 28), если не однозначно показывают близкое генетическое сходство между калмыками, бурятами и монголами Монголии и Внутренней Монголии (АРВМ КНР), то, по крайней мере, позволяют очертить выраженные скопления монголоязычных групп. Начиная с 3-х систем (рис. 4), с появлением в анализе данных по системе RH, популяции начинают явно разделяться на европеоидные и монголоидные, т.е. результаты для территории Северной Евразии отражают сильный географический клин генетических частот популяций в направлении восток-запад.
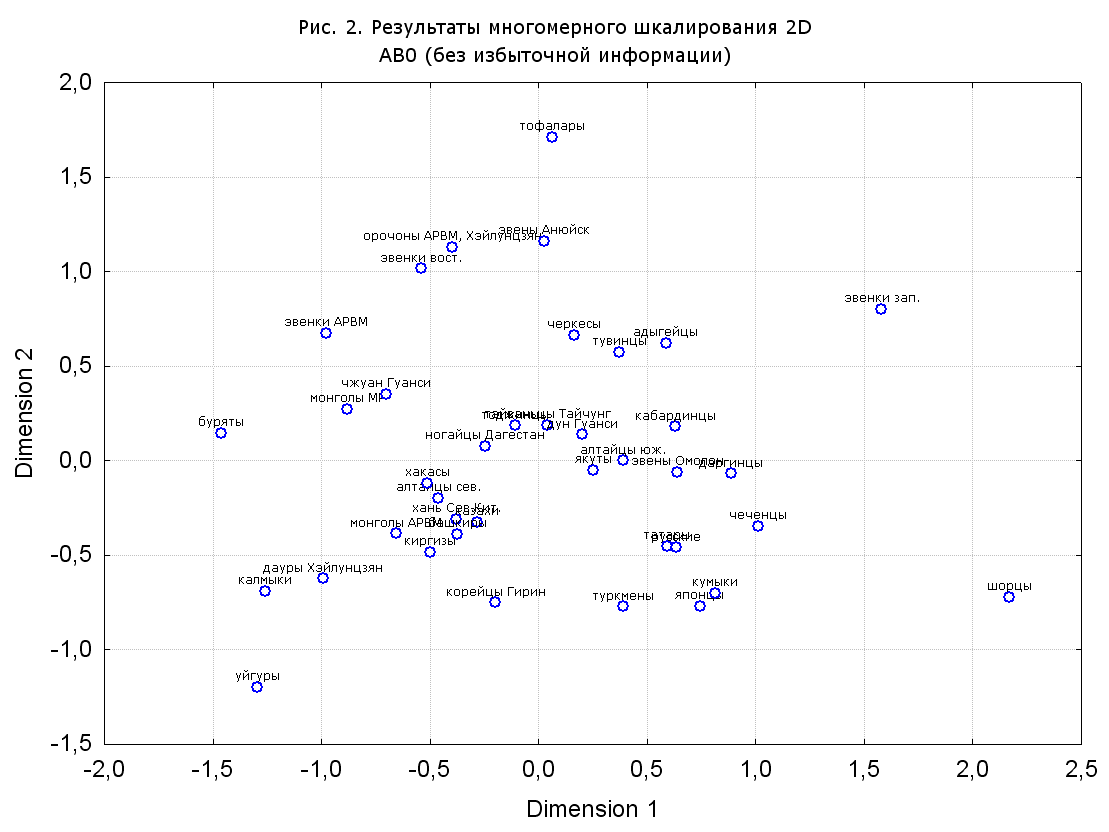
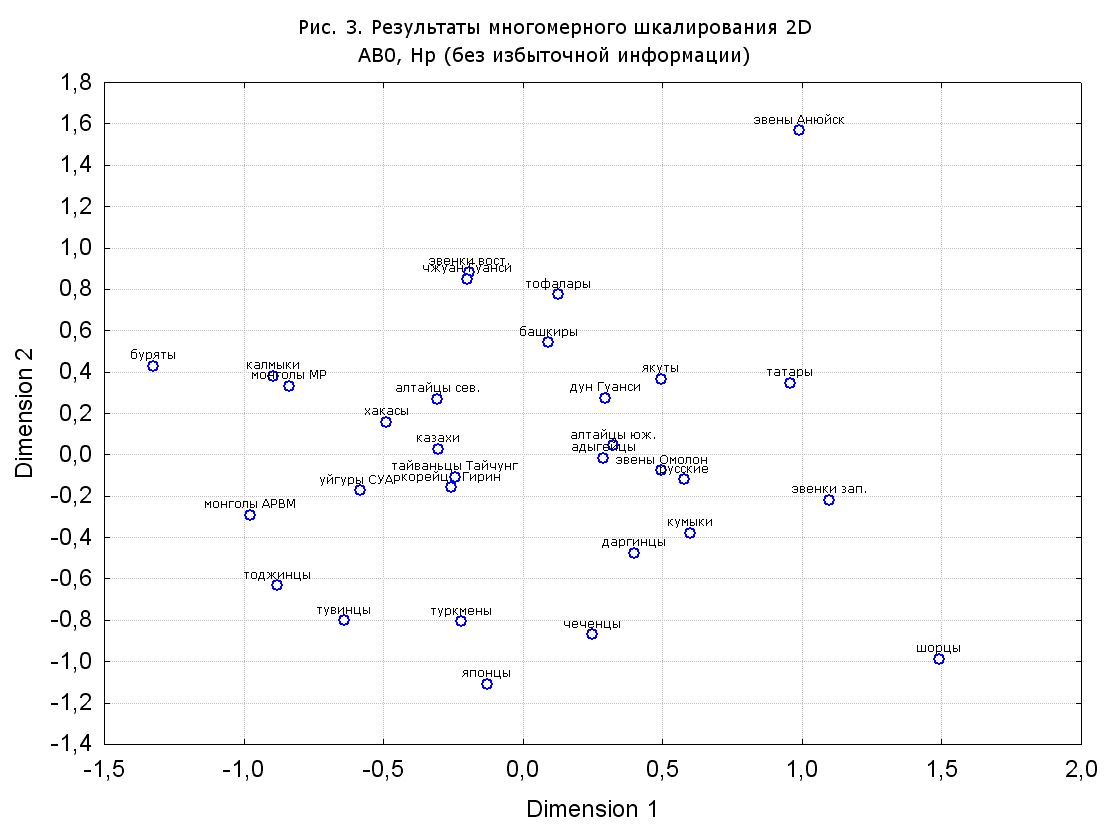
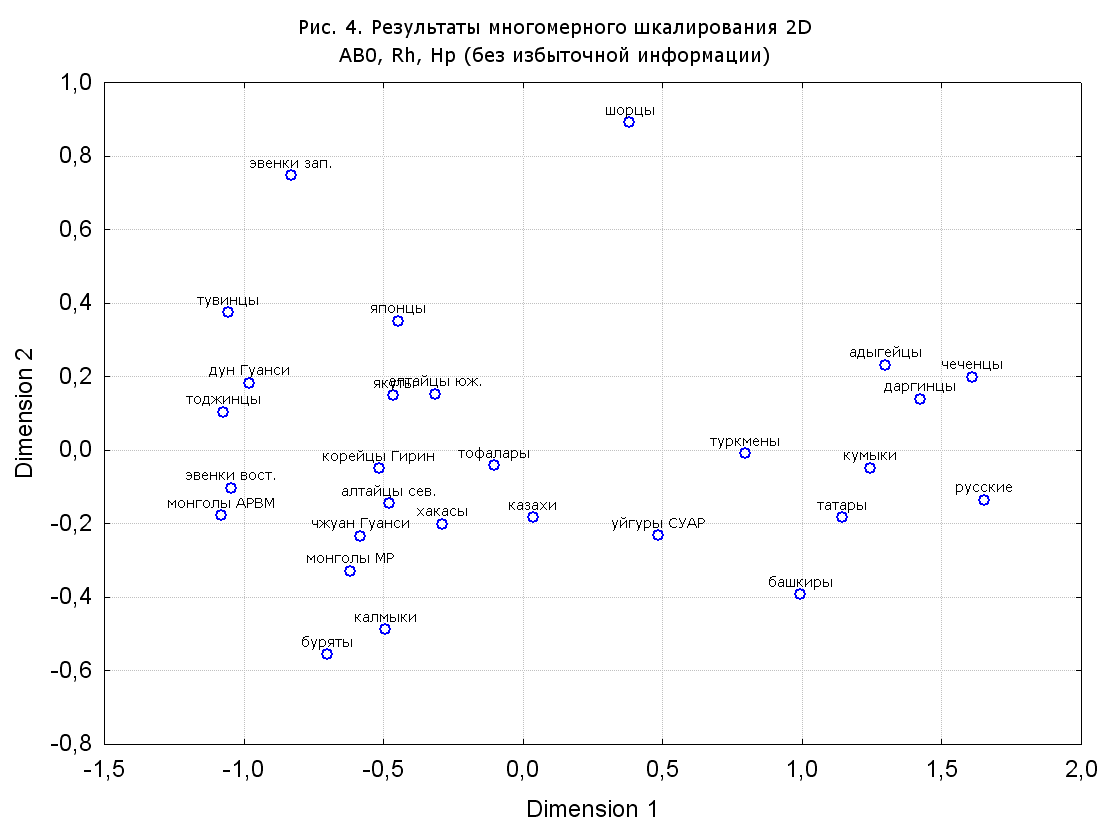
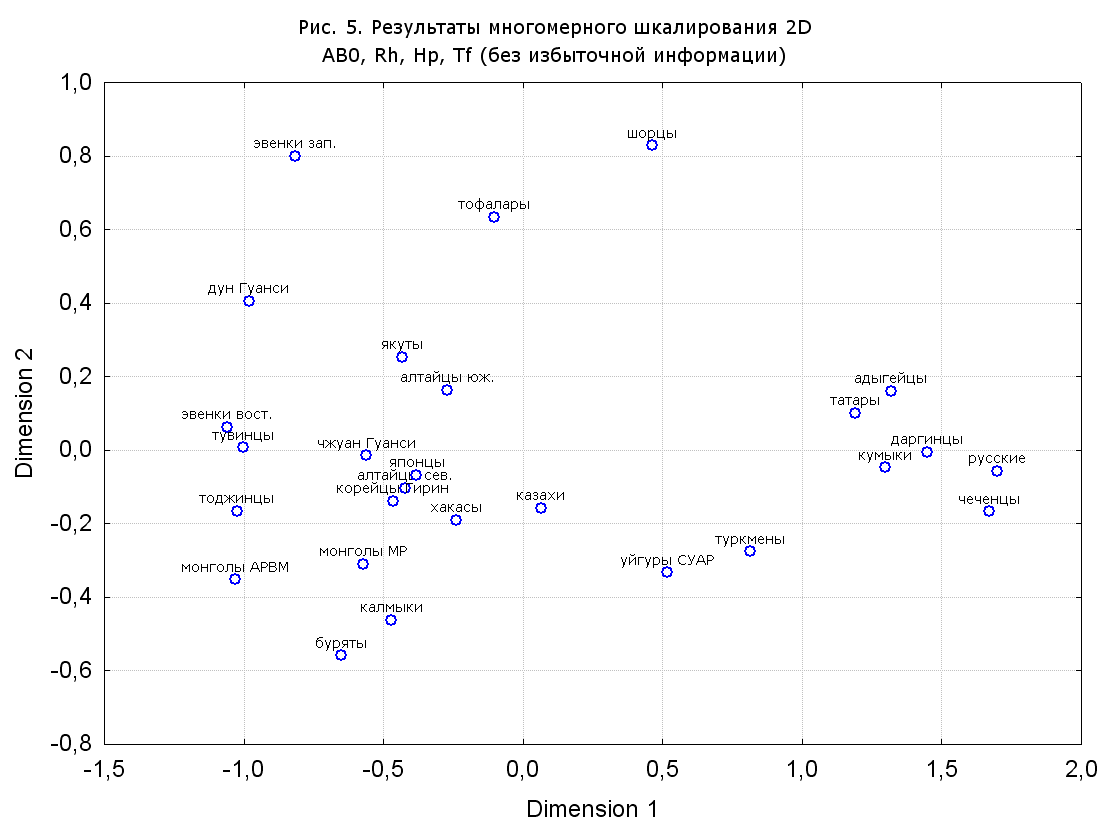
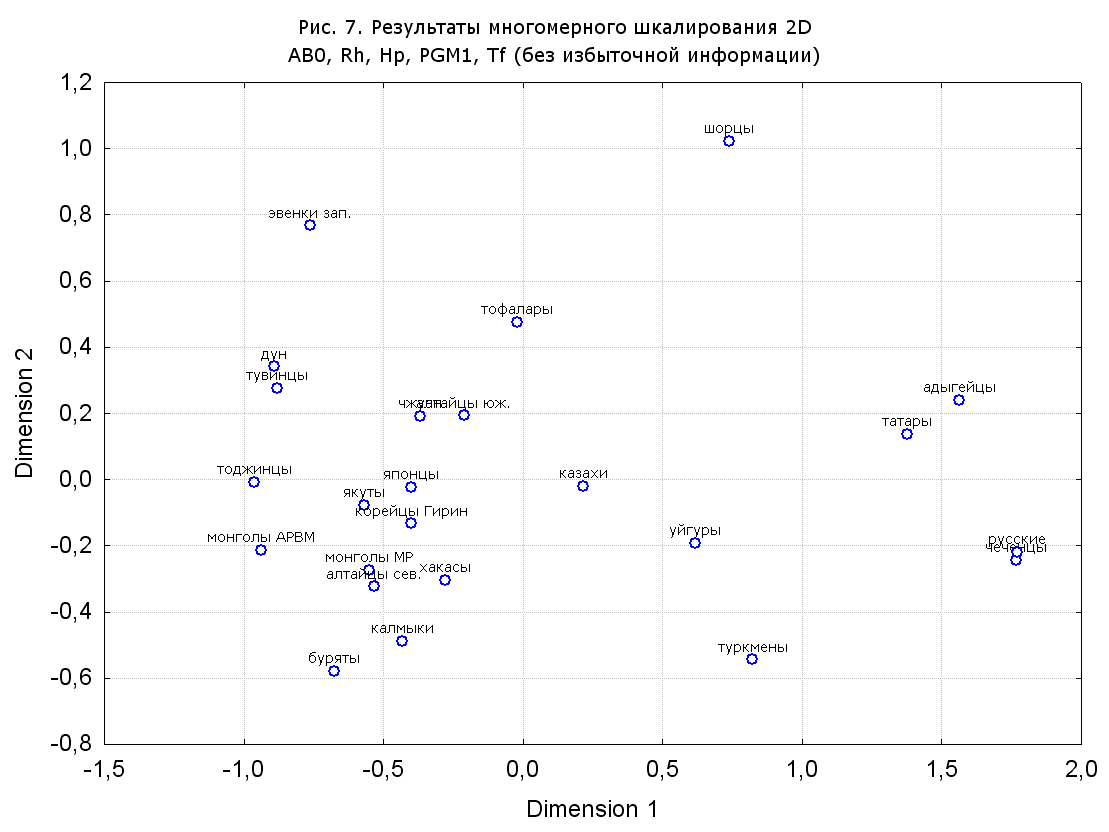
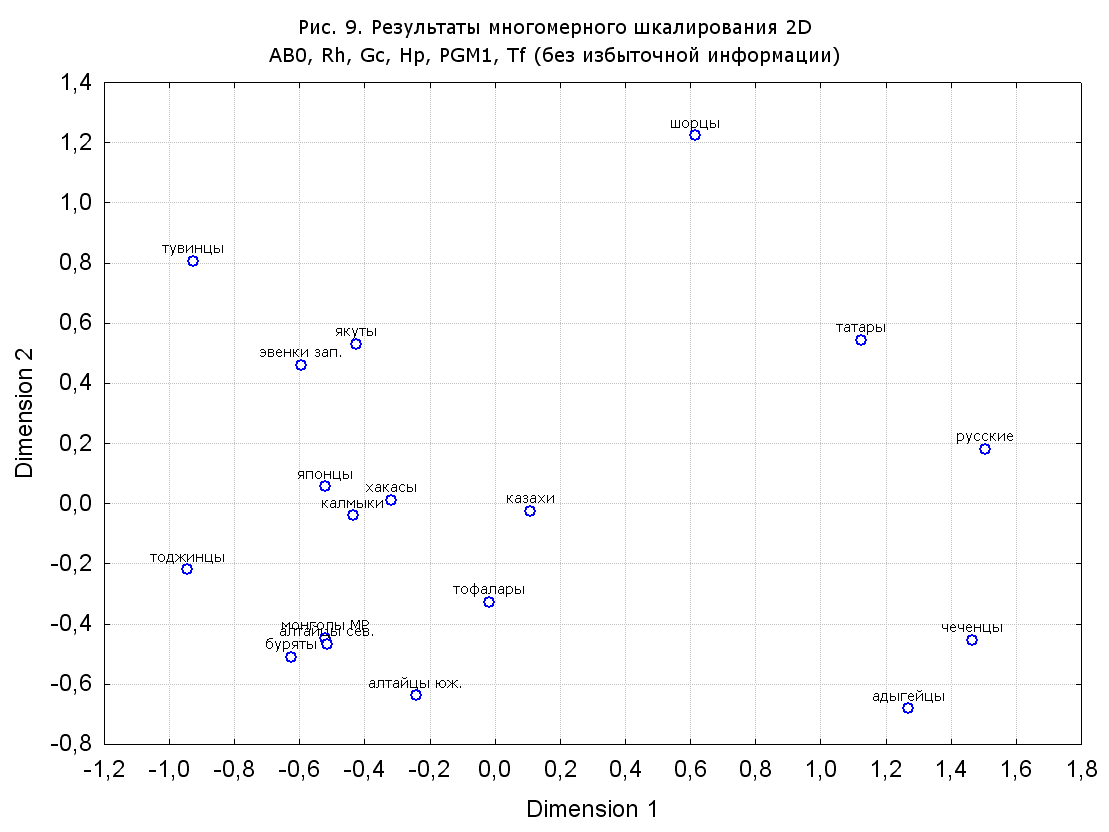
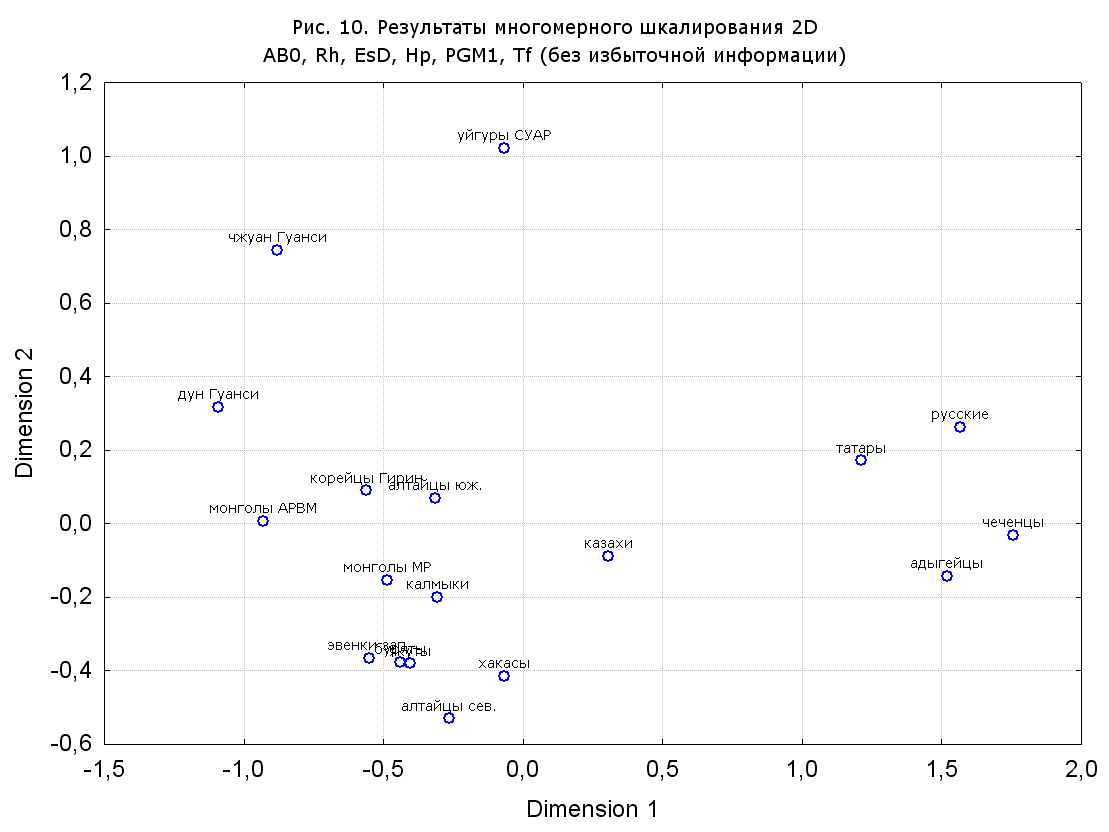
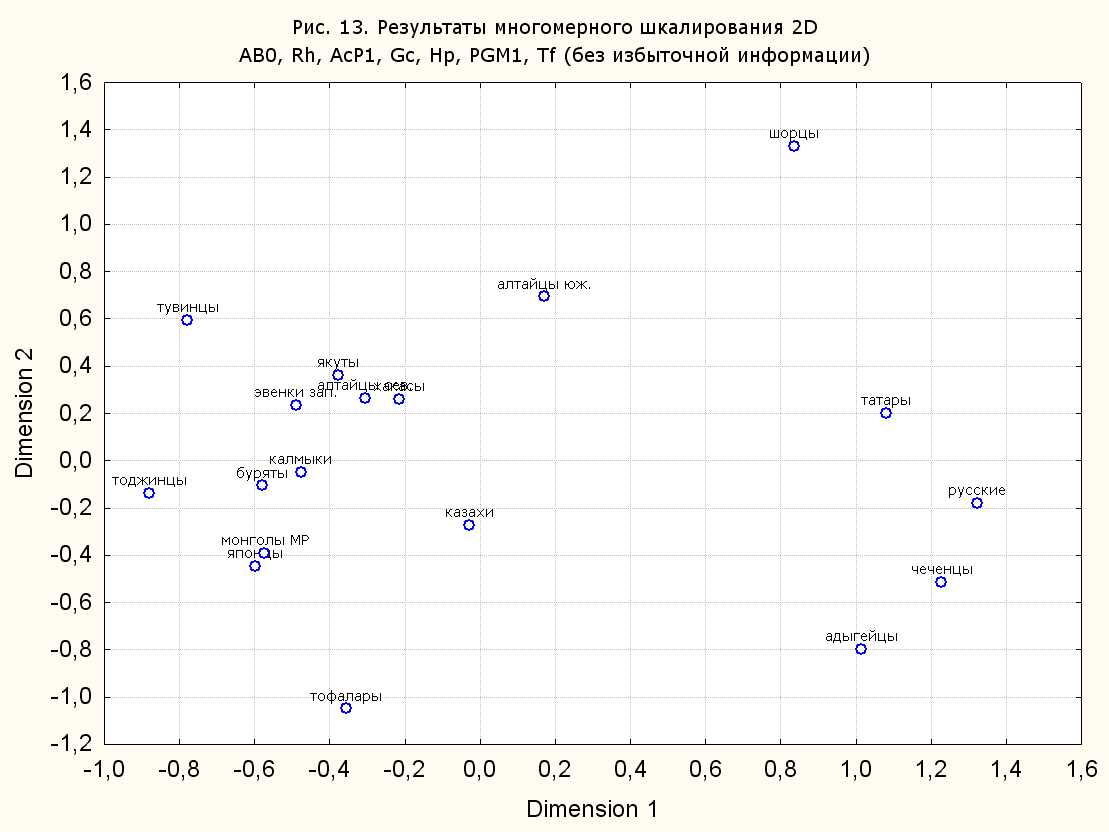
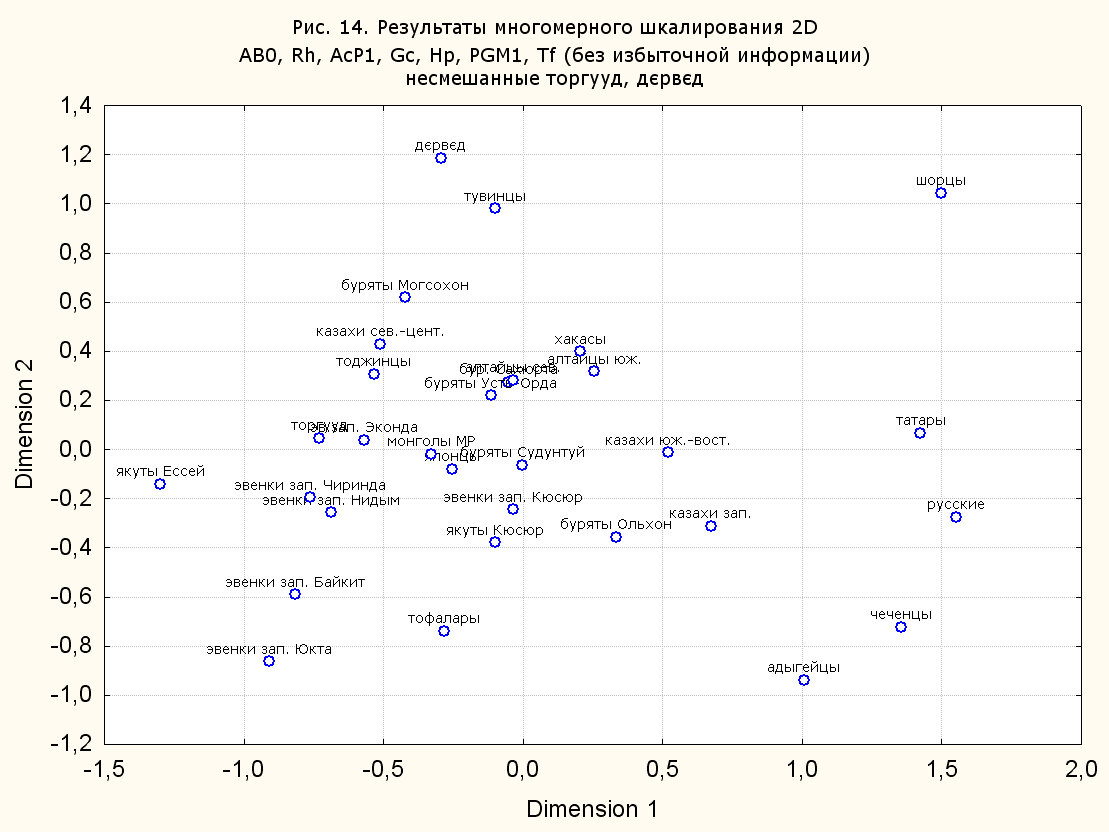
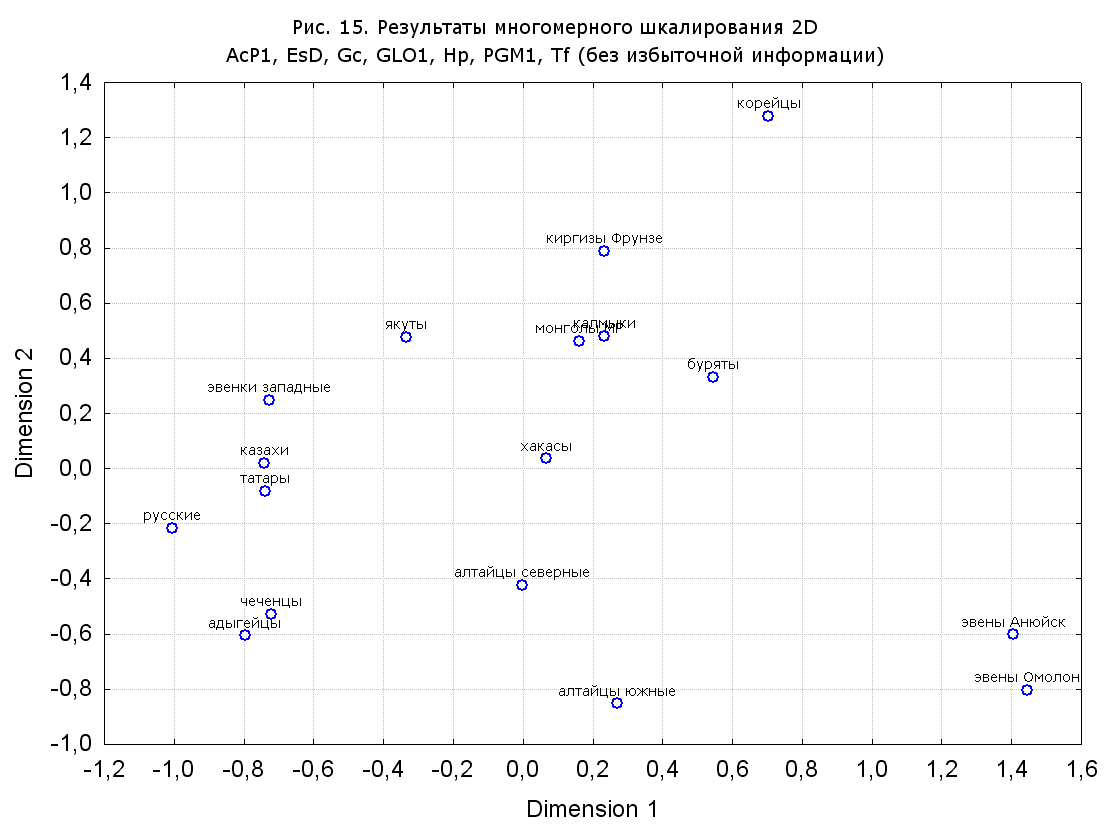
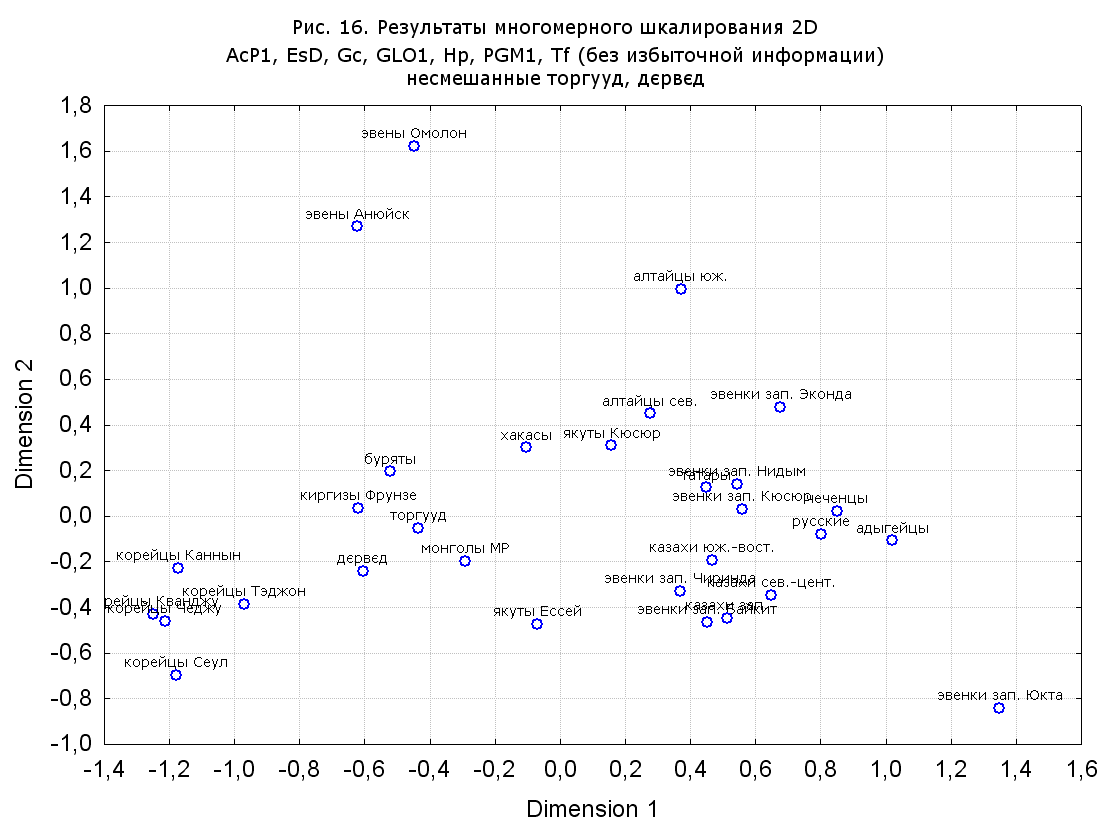
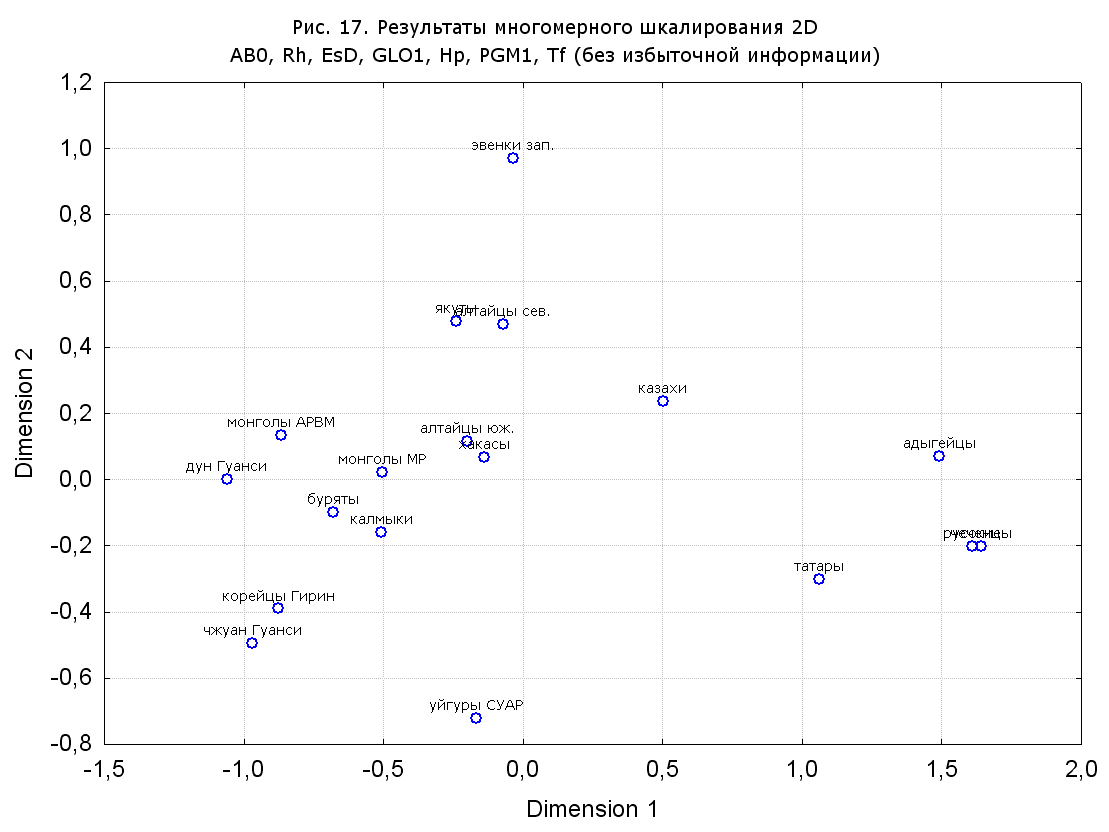
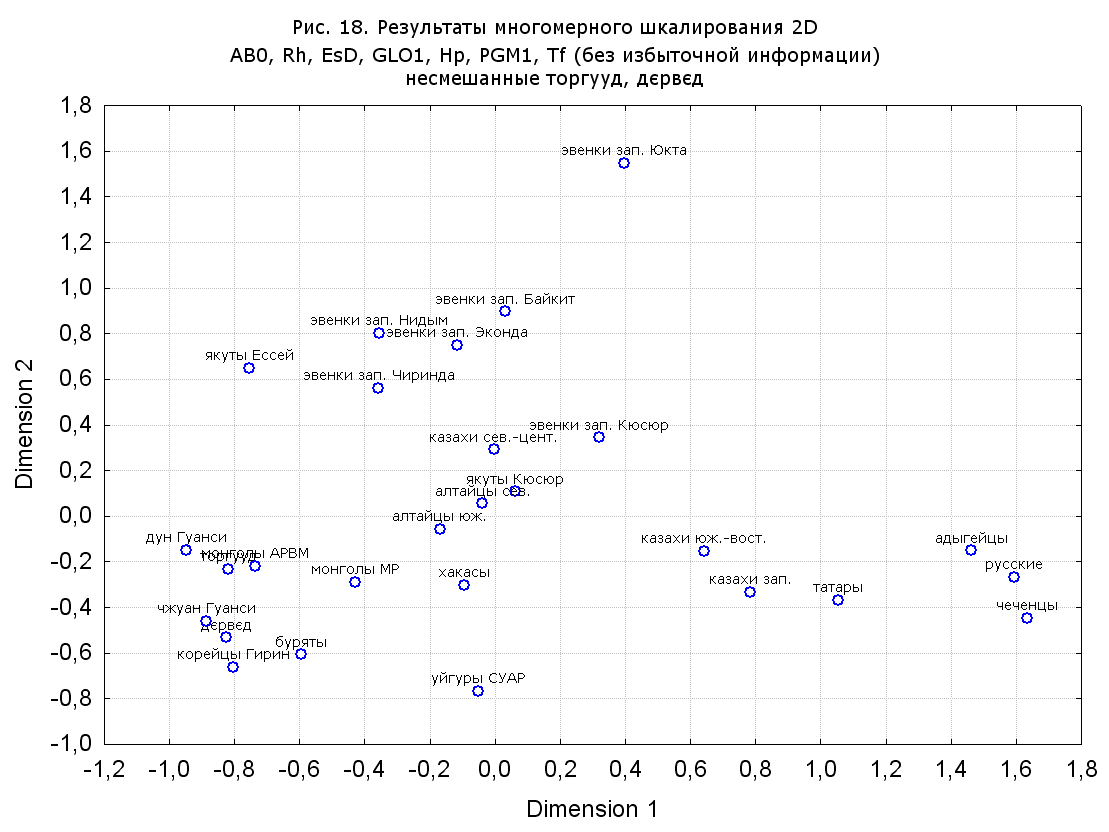
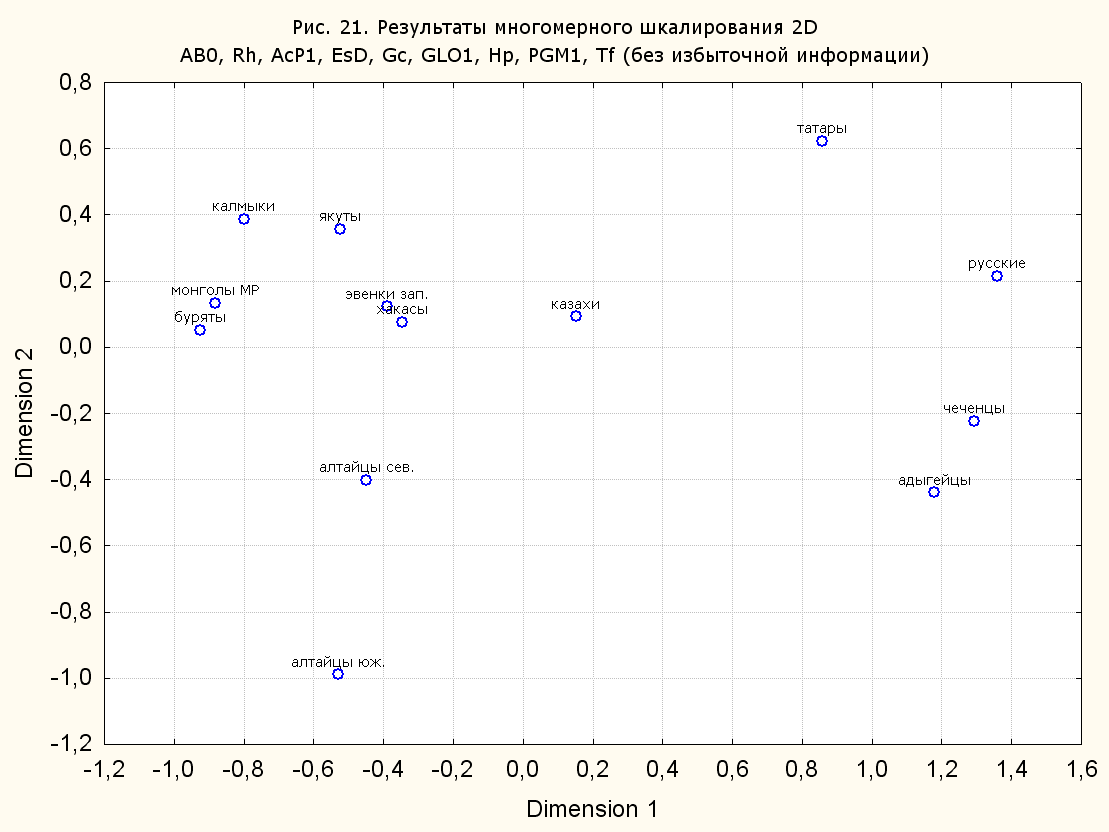
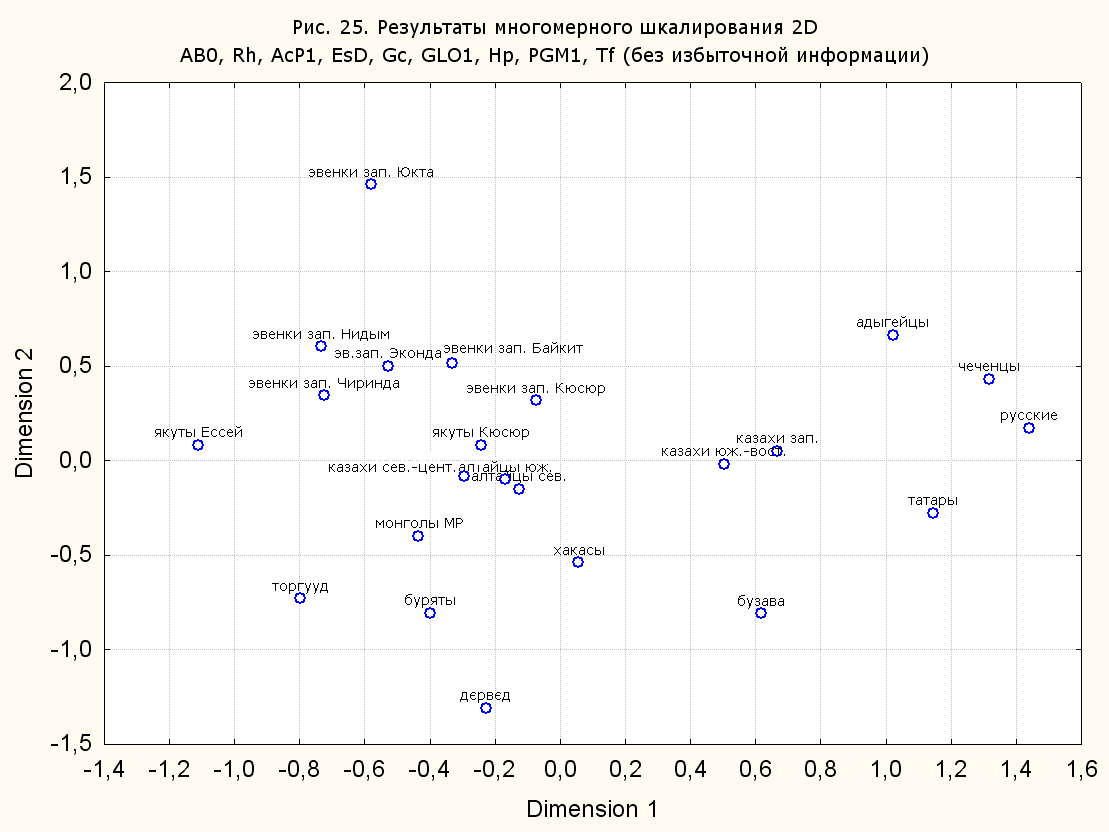
Значимость системы RH подчеркивается результатами шкалирования данных по 7 системам (рис. 13-18). При исключении из анализа частот по данной системе (рис. 15-16) генетические расстояния между монголоидными группами начинают превышать расстояния между монголоидными и европеоидными.
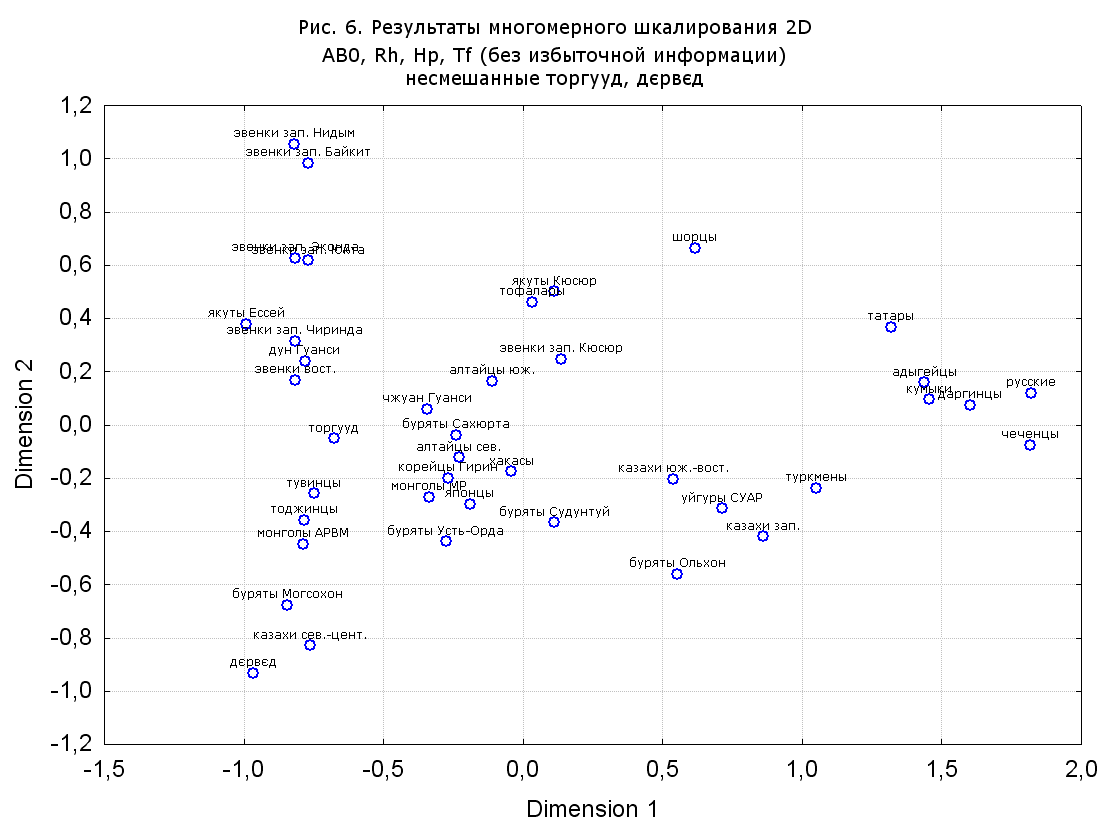
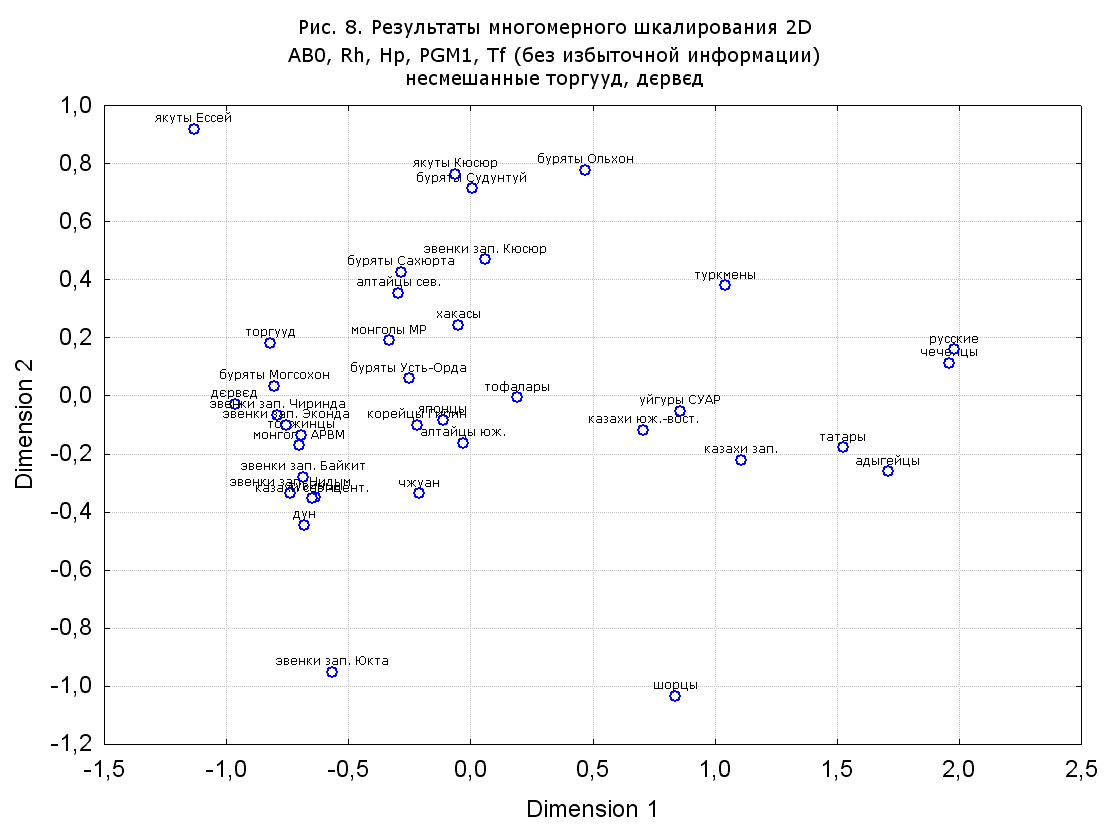
При разбиении данных по калмыкам в целом на данные по торгууд, дєрвєд и бузава (рис. 6, 9, 10, 13, 15, 17, 20, 29), результаты многомерного шкалирования показывают вполне достоверные отличия этих групп. В большинстве случаев, в том числе и не показанных в данной статье, генетические расстояния между бузава и дєрвєд гораздо ближе друг другу, нежели к торгууд (рис. 1, 26, 27). При этом частоты генов бузава в целом (рис. 24, 25) более смещены к европеоидным группам, нежели частоты несмешанных бузава (рис. 26, 27). Близость бузава и дєрвєд, скорее всего, объясняется тем, что бузава, попавшие в нашу выборку, большей частью являются выходцами из дєрвєдских аймагов, нежели из торгуудских. Значительная разница между частотами генов бузава в целом и несмешанных бузава возникла, видимо, в силу того, что 2 индивидуума, попавшие в первую группу, имели в своей родословной русскую прапрабабку, что по всей вероятности сильно отразилось на частотах генов небольшой выборки (19 чел.).
Положение дєрвєд хотя и характеризуется близостью монголоидным группам, но в большинстве случаев довольно обособлено от большей их части. Относительно торгууд это положение имеет некоторую тенденцию смещения в сторону европеоидных групп (рис. 19, 22-27). Генетические расстояния, вычисленные на основании частот разных наборов систем генетических маркеров, чаще показывают в ближайших соседях дєрвєд казахов Северо-Центральной этноисторической области, бурят с. Могсохон, тувинцев (рис. 6, 8, 11, 14, 19).
Положение генетических частот торгууд даже «монголоиднее» средних частот генов по монголам Монголии и ближе им относительно дєрвєд. Кроме того, относительно дєрвєд, в ряде случаев торгууд обнаруживают если не близость различным эвенкийским группам, то тенденцию сближения с ними. Результаты многомерного шкалирования на основании частот разных наборов систем генетических маркеров чаще всего показывают близость торгууд монголам МР и АРВМ КНР, эвенкам, бурятам и в паре случаев дун из пров. Гуанси КНР (рис. 6, 8, 11, 12, 14, 18, 19, 22, 23).
Обсуждение
Первым и довольно ожидаемым результатом нашего исследования является констатация факта близости калмыков в целом азиатским популяциям, и, в частности, монголоязычным группам – монголам Монголии и бурят-монголам РФ. Учитывая исторические факты, лингвистические и этнографические сведения, ничего удивительного в этом факте нет. Ойраты (или ойрат-монголы) наряду с другими монголоязычными группами являются частью монгольской этнической общности. Учитывая несколько сот лет, прошедших для европейской ветви ойрат в иноэтничном окружении, в изоляции от родственных групп, можно было бы ожидать сдвига аллельных частот в сторону европеоидных групп. Однако на нашем материале это предположение в основном не подтверждается, и в этом полученные нами результаты сходятся с данными, полученными классическими антропологическими методами [Чебоксаров 1935; Ашилова 1976].
Если рассматривать отдельные калмыцкие группы, то, несмотря на предполагаемую возможность их смешения между собой, картина генетических расстояний показывает достоверные различия между ними. При этом относительно других групп частоты генов торгууд показывают наибольшую близость усредненным частотам генов по монголам Монголии, а по оси «монголоидность-европеоидность» даже «монголоиднее» их. Данные по бузава можно не учитывать, поскольку выборка бузава не достигает статистически значимой величины. Следует отметить, что даже если бы она была статистически значимой, то, учитывая факт сложения бузава из торгууд, дєрвєд и зvvнгар, близость бузава к дєрвєд не означает близости вообще всех бузава, а может толковаться как близость к ним большинства индивидуумов, попавших в нашу в выборку.
Отметим, что достоверное различие современных европейских торгууд и дєрвєд с несколько более выраженным смещением торгууд в сторону монголоидных, а дєрвєд – в сторону европеоидных групп, впервые показанное нами с помощью генетических маркеров белковых систем [Galushkin 1998; Galushkin et al. 2001] и мт-ДНК [Ovtchinnikova et al. 1997], фиксировалось ранее и на основе сомато- и кефалометрии. Чебоксаров [1935] и Ашилова [1976] отмечали ослабление комплекса монголоидных черт среди некоторых калмыцких групп. Одним из возможных объяснений заметных морфологических различий между калмыцкими группами является некоторое смешение с окружающими, большей частью европеоидными популяциями, среди которых в недавнем прошлом преобладали тюркоязычные группы. Взаимодействия с татарами, туркменами, адыгами, кумыками, карачаевцами и другими этническими группами могли способствовать появлению европеоидных расовых типов в структуре европейских ойрат, по крайней мере, в течение последних 250-300 лет [Чебоксаров 1935]. Однако фиксируемые морфологические и генетические различия не обязательно являются результатом последних сотен лет. Вполне возможно, эти различия существовали еще до прихода ойратских групп на европейские территории. Они могли обуславливаться более ранними контактами с разными народами, интенсивность и длительность которых была неодинакова для разных ойратских групп. Кроме этого, следует учитывать тот факт, что само сложение этих групп могло идти на основе субстрата, исходно имевшего различные соотношения антропологических типов и генетических линий. Для выявления вклада в фиксируемые нами различия каждого из периодов, необходим детальный исторический анализ, которого мы, к сожалению, не имеем до сих пор.
Необходимо отметить еще один момент. Морфологические и генетические различия фиксируются у современного населения, несмотря на то, что ойраты являлись союзом, внутри которого происходили перемещения и обмен группами населения. Сохранению и, вероятно, накоплению различий между ойратскими группами мог способствовать тот факт, что, будучи объединенными политически, они не были и не успели стать гомогенными генетически, имея, по всей видимости, различные истории формирования.
Несмотря на констатацию морфологических и генетических различий современных европейских ойратских групп, никем из генетиков, историков и прочих специалистов, занимавшихся вопросами ойратского (в том числе калмыцкого) этногенеза попыток объяснить эти различия не делалось. Отдельно стоит упомянуть работы Н.Н. Чебоксарова [1935] и В.П. Алексеева [1980]. Н.Н. Чебоксаров на основании кефалометрии дєрвєд и бузава относил появление европеоидных признаков у западных калмыков к европейскому периоду их истории, а формирование различных монголоидных признаков – к периоду пребывания ойрат в Азии. В.П. Алексеев, на основании изучения сборной выборки калмыцких черепов неясной этнотерриториальной принадлежности, приходил к выводу о том, что европеоидный компонент восходит в своём происхождении к азиатскому периоду истории ойрат. В отличие от выводов на основе морфометрического материала, интерпретация генетических результатов может осложняться тем, что морфологические черты, будучи более экочувствительны, не обязательно могут изменяться согласованно с генетическими [Galushkin et al. 2001]. Это подтверждается, например, на популяциях менонитов, разошедшихся более 100 лет назад, у которых морфологическое и генетическое сходство несогласованно [Crawford et al. 2000]. Мы понимаем, насколько трудно в наши дни говорить о периоде возникновения различных европеоидных и монголоидных элементов и генетических черт в составе современных ойратских групп, однако некоторые имеющиеся в нашем распоряжении факты позволяют сделать определенные предположения.
Данные языкознания свидетельствуют о том, что основными диалектами современного языка европейских ойрат, имеющего в некоторых классификациях название калмыцкого, являются диалекты дєрвєд и торгууд. Стремление к редукции гласных и более твердый выговор у дєрвєд в отличие от торгууд являются одними из многих черт, разделяющих эти диалекты. Указанные черты могут являться результатом влияния тюркских языков. К сожалению, нам не встречались работы, в которых бы специально анализировалось влияние тех или иных тюркских языков на различные ойратские диалекты. Однако, если наше предположение верно, то следует заметить, что современные представители различных тюркоязычных групп, с предками которых ойраты контактировали в период средневековья, не о говоря уже о древности, в массе своей заметно европеоиднее современных представителей монголоязычных народов. Собственно говоря, этот факт известен и безотносительно лингвистических сведений. Таким образом, бо’льшая морфологическая европеоидность современных дєрвєд относительно торгууд и их генетические различия могут объясняться более длительными и/или интенсивными контактами предков дєрвєд с группами, имевшими бо’льшую долю европеоидности и являвшихся носителями тюркских языков. Для установления периодов и ареалов этих контактов требуется отдельное историческое исследование. Кроме этого, не исключено, что само сложение дєрвєд изначально шло на основе более европеоидных, относительно предков торгууд, групп населения. Результаты, представленные в данной работе показывают, что в некоторых случаях наиболее близкими современным европейским дєрвєд оказывается часть казахов, буряты и тувинцы. Однако для отслеживания общих предков этих групп требуется проделать немалое историческое исследование, честь проведения которого мы оставляем для других исследователей.
Что касается торгууд, то наиболее популярная версия их происхождения гласит, что эта этническая группа ведет свое начало от охранного корпуса Чингис-хана tur[g]aq kesikten ([g] в данном случае читать как гамму) [Авляев, Санчиров 1984; Санчиров 1996]. Ранее в свете этой гипотезы нами было высказано предположение, что бо’льшая монголоидность торгууд относительно дєрвєд по морфологическим и генетическим системам признаков, может объясняться, по всей видимости, тем, что в XIII-XIV вв. охранный корпус был дислоцирован в Китае. Известно, что в 1260 г. Хубилай перенес монгольскую столицу в Кайпин (Шанду, Шангду), в 1264 г. в Яньцзин (Пекин), а в 1271-1272 гг. в Хаан Балгасун (Дай-ду) [Далай 1983: 43], конец же монгольскому владению Китаем наступил после поражения 1388 г. Неизвестно, могла ли гвардия императора за почти двухсотлетний период измениться генетически, путем смешения с окружающим населением, но вероятность подобного события нельзя не сбрасывать со счета [Хойт 2005]. В пользу этого предположения могут говорить зафиксированные Дарваевым [2003] особенности современного торгуудского говора, который «характеризуется бо’льшим консерватизмом и близостью к исторической основе, чем другие говоры калмыцкого языка», при этом «мелодическая подвижность речи торгутов - это отражение влияния китайской речи, где высота тона играет смыслоразличительную роль». С другой стороны, известно, что социальное положение китайцев в монгольской империи было низким, и этот факт говорит не в пользу того, что возможное потомство от предполагаемых контактов монгольских солдат с китайскими женщинами могло поступить в армию, и тем более в элитные гвардейские подразделения.
В связи с этим встает ряд вопросов о способе пополнения и набора в гвардию, особенно учитывая постоянно ведущиеся военные действия, при котором средний срок жизни солдата в монгольской империи не мог быть большим. Каков был этнический состав гвардии? Набиралась ли гвардия по мере необходимости из различных или только определенных монгольских улусов, был ли это наследственный институт, куда шли служить дети гвардейцев или это было и то и другое вместе? Могли ли находиться при ставке семьи гвардейцев или они обзаводились ими у себя на родине? Какова была вероятность смешанных браков и дальнейшей службы в гвардии потомства от этих браков? Данные, лежащие на поверхности, вопросы историками до сих пор не освещены, и потому гипотеза о возможном влиянии китайского населения XIII в., на генетическое положение современных торгууд РК РФ остается открытой.
Гипотезу о китайском влиянии можно отбросить, если рассматривать более ранний, доимперский период истории монгольских народов. Согласно Грум-Гржимайло [1926], заселение высокогорного степного плато, более или менее соответствующего территории современной Монголии, после падения Уйгурского государства в 840 г. шло с востока и осуществлялось группами, известными под общим этнонимом татар. В XII в. они стали доминировать не только на территории современной Восточной Монголии, после чего имя татар распространилось на всех, или, по крайней мере, на большинство кочевников [Гумилев 1970]. Этноним татар в данный период являлся, по всей видимости, тюркским наименованием групп, в китайской историографии именуемых шивэй [Васильев 1997]. В свою очередь, под наименованием шивэй могли фигурировать различные в лингвистическом плане этнические группы, сведениями об антропологическом облике которых мы не располагаем. Известно, что современными потомками шивэй является ряд монголо- и тунгусо-маньчжуроязычных народов. В свете этих данных более «монголоидное» положение торгууд относительно дєрвєд, их бо’льшая близость различным монгольским группам и эвенкам, которые мы наблюдаем на картах генетических расстояний, может говорить не о китайском влиянии, как мы предполагали ранее, а о дальневосточном субстрате, послужившем основой для современных торгууд в бо’льшей степени нежели для современных дєрвєд.
В заключение, как и четыре года назад [Хойт 2005], отметим, что для проверки результатов нашего популяционно-генетического исследования необходимо провести аналогичную работу по ойратским группам Монголии и Китая. Добавим при этом, что для решения как частных (сложение отдельных групп), так и общих проблем ойратского этногенеза желательно привлечь весь корпус имеющихся исторических, этнологических и лингвистических данных, а при наличии пробелов в этих данных, провести специальные исследования.
ЛИТЕРАТУРА
Авляев Г.О., Санчиров В.П. К вопросу о происхождении торгоутов и хошоутов в этническом составе средневековых ойратов Джунгарии (к проблеме этногенеза калмыков) // Проблемы этногенеза калмыков. Элиста, 1984.
Алексеев В.П. К краниологии калмыков в связи с их происхождением // Вопросы сравнительной этнографии и антропологии калмыков. Элиста, 1980. C. 3-41.
Алексеева Т.И., Волков-Дубровин В.П., Павловский О.М., Смирнова Н.С., Спицын В.А., Щекочихина Л.К. Антропологические исследования в Забайкалье в связи с проблемой адаптации у человека (морфология, физиология и популяционная генетика). I. Население Баргузинской котловины в свете исторических и популяционно-генетических данных // Вопросы антропологии. 1970. Вып. 36. C. 3-19.
Ашилова Д.О. Этническая антропология калмыков: Соматологическое исследование. Элиста: Калм. кн. изд-во, 1976. – 215 с.
Батсуурь Ж., Самбуугийн Н., Шнейдер Ю.В., Петрищев В.Н., Рычков Ю.Г. Генетическая структура монголов по локусам АВ0, MN, Rh, EsD, GLO1, PGM1, AcP, 6-PGD, Hp, Gc, Tf, C'3 и ChE2 // Генетика. 1991. Т. 27. № 2. С. 316-326.
Васильев А.А. Глава IV. Монголы и монгольское завоевание. Монголия в первой половине XII в. // История Востока. Восток в Средние века. М.: «Восточная литература» РАН, 1997.
Генофонд и геногеография народонаселения / Под ред. Ю.Г. Рычкова: Том 1. Генофонд населения России и сопредельных стран. СПб.: Наука, 2000. – 611 с.
Грум-Гржимайло Г.Е. Западная Монголия и Урянхайский край: В 3-х т. Т. II. Л., 1926.
Гумилев Л.Н. Поиски вымышленного царства. М., 1970.
Далай Ч. Монголия в XIII-XIV веках. Пер. с монг. П. Нихлая. М.: Наука, 1983. – 232 с.
Дарваев П.А. Калмыцкий язык в свете теории культуры языка и речи. Монография. Элиста: АПП «Джангар», 2003. – 280 с.
Дейвисон М. Многомерное шкалирование. Москва: Финансы и статистика, 1988.
Животовский Л.А. Популяционная биометрия М.: Наука, 1991. – 271 с.
Левин М.Г., Трофимова Т.А. Калмыки. Краниологический очерк // Антропологический журнал. 1937. № 1. C. 73-81.
Санчиров В.П. О Происхождении этнонима торгут и народа, носившего это название // Монголо-бурятские этнонимы: Сб. ст. Улан-Удэ: БНЦ СО РАН, 1996. С. 31-50.
Сокровенное сказание монголов: Анонимная монгольская хроника 1240 г. Элиста: Калмыцкое книжное издательство, 1990. – 280 с.
Спицын В.А. Биохимический полиморфизм человека. М.: Изд-во Моск. ун-та, 1985. – 216 с.
Спицын В.А., Афанасьева И.С., Агапова Р.К., Цурикова Г.В., Щекотихина Ю.А., Краузе Д., Куххайзер В. Изучение генетических маркеров у русских и немцев в рамках совместного Российско-Германского исследовательского проекта // Генетика. 1994. Т. 30. № 5. С. 702-708.
Спицын В.А., Титенко Н.В. Подтипы группоспецифического компонента (GC) в норме и при некоторых заболеваниях // Генетика. 1990. Т. 26. С. 749-759.
Терехина А.Ю. Анализ данных методами многомерного шкалирования. Москва: Наука, 1986.
Фогель Ф., Мотульски А. Генетика человека: В 3-х т. Том 2: Пер с англ.. М.: Мир, 1990. – 378 с.
Хойт С.К. Неясные моменты этногенеза монголоязычных групп на примере торгутов РК РФ // Трофим Алексеевич Бертагаев: к 100-летию со дня рождения. Сб. статей. Элиста: ЗАОрНПП «Джангар», 2005. С. 330-336.
Хойт С.К. Последние данные по локализации и численности ойрат // Проблемы этногенеза и этнической культуры тюрко-монгольских народов. Вып. 2. Элиста: Изд-во КГУ, 2008. С. 136-157.
Чебоксаров Н.Н. Калмыки Западного улуса. Расово-антропологический очерк // Антропологический журнал. 1935. №1. С. 21-62.
Шилэгийн Норбо. Зая-пандита (материалы к биографии). Пер. со старомонг. Д.Н. Музраевой, В.П. Санчирова, К.В. Орловой. Элиста: Калм. кн. изд-во, 1999. – 335 с.
Эрдниев У.Э. Калмыки. Элиста: Калм. кн. изд-во, 1985. – 282 с.
Benkmann H.G., Cho Y.H., Singh S., Wimmer U., Lee C.C., Kim I.K., Paik Y.K., Goedde H.W. Red Cell Enzyme and Serum Protein Polymorphisms in South Korea // Human Heredity. 1989. № 39. P. 263-270.
Crawford M.H., Demarchi D., Ellerd et al. Body morphology and aging among the Mennonites of Kansas and Nebraska // Different Seasons. Biological Aging among the Mennonites of Midwestern United States. University of Kansas Publications in Anthropology, 21. Lawrence, KS, 2000. P. 55-67.
Crawford M.H., McComb J., Schanfield M.S. et al. Genetic structure of pastopal populations of Siberia: The Evenki of Central Siberia and the Kizhi of Gorno Altai // The Last of the Pastoralists: Biocultural Dimentions, Leonard W. and Crawford M.H., eds. Cambridge, UK: Cambridge University Press, 2001.
Divall G.B. The ESD polymorphism as revealed by IEF in ultra-thin polyacrylamide gels // Forensic Sci. Int. 1984. № 26. P. 255-267.
Du Ruo-fu and Chen Liang-zhong. A Preliminary Study on Genetic Polymorphism of Chinese Nationalities // Chinese-American Symposium. 1986. P. 134-155.
Galushkin S.K. State of Kalmyk groups among some populations of Eurasia // 11th Congress of European Anthropological Association "Humans and Environment". 30 August - 3 September 1998. Program Abstracts. P. 91-92.
Galushkin S.K., Spitsyn V.A., Crawford M.H. Genetic Structure of Mongolic-speaking Kalmyks // Human biology. December 2001. Vol. 73. № 6. P. 823-834.
Goedde H-W., Benkman H-G. and Hirth L. 1981 Ultrathin-layer isoelectric focusing of subtypes of GC, TF, Pi and PGM1 // Biomathematical Evidence of Paternity, К. Hummele and J. Gerchow, eds. Berlin: Springer-Verlag, 1981. P. 123-126.
Gorg A., Weser J., Westermeier R., Postel W., Weidinger S., Patutschnick W., Cleve H. Isoelectric focusing with immobilized pH gradients for the analysis of transferrin (Tf) subtypes and variants // Human Genetics. 1983. V. 64. P. 222-226.
Jing Y. Lin. The Distribution of the Polymorphic Groups of Blood, Serum Protein and Red Cell Enzime of the Taiwanese // J. Anthropol. Soc. Nippon. 1975. 83, № 2. P. 203-211.
Miller S.A., Neelson M.S., Dykes D.D., et al. 1987 Comparison of acid phosphatase ACPl variants by IEF and conventional electrophoresis: Identification of three new alleles, ACP*N, ACP1*P, and ACP1*S // Human Heredity 1987. № 37. P. 371-375.
Nei M. Genetic distance between populations // Amer. Natur. 1972. Vol. 106. P. 283-292.
Novoradovsky A.G., Spitsyn V.A., Duggirala R., Crawford M.H. Population genetics and structure of Buryats from the Lake Baikal Region of Siberia // Human Biology. 1993. Oct. № 65(5). P. 689-710.
Omoto K., Ishizaki K., Harada S., Akaishi S., Kudo T. and Takahashi K. The distribution of serum protein and red cell enzyme types among blood donors of Okinawa Island, the Ryukyus // Journal of the Anthropological Society of Nippon, 1973. № 81. P. 159-173.
Ovtchinnikova O., Druzina E., Galushkin S., Spitsyn V.A., Ovchinnikov I.V. An Asian-specific 9-bp deletion in region V of mitochondrial DNA is found in Europe // 9. Jahrestagung der Gesellschaft für Humangenetik. Medizinische Genetik. 1997. №1. P. 85.
предыдущая | в содержание
to the library | номын сан руу | в библиотеку